UNITED STATES
SECURITIES AND EXCHANGE COMMISSION
Washington, D.C. 20549
FORM
(Mark One)
|
|
ANNUAL REPORT PURSUANT TO SECTION 13 OR 15(d) OF THE SECURITIES EXCHANGE ACT OF 1934 |
For the fiscal year ended
OR
|
|
TRANSITION REPORT PURSUANT TO SECTION 13 OR 15(d) OF THE SECURITIES EXCHANGE ACT OF 1934 FOR THE TRANSITION PERIOD FROM TO |
Commission File Number
(Exact name of Registrant as specified in its Charter)
|
|
|
|
(State or other jurisdiction of incorporation or organization) |
(I.R.S. Employer Identification No.) |
|
|
|
|
(Address of principal executive offices) |
(Zip Code) |
Registrant’s telephone number, including area code: (
Securities registered pursuant to Section 12(b) of the Act:
|
Title of each class |
|
Trading Symbol(s) |
|
Name of each exchange on which registered |
|
|
|
|
|
|
Securities registered pursuant to Section 12(g) of the Act: None
Indicate by check mark if the Registrant is a well-known seasoned issuer, as defined in Rule 405 of the Securities Act.
Indicate by check mark if the Registrant is not required to file reports pursuant to Section 13 or 15(d) of the Act. Yes ☐
Indicate by check mark whether the Registrant: (1) has filed all reports required to be filed by Section 13 or 15(d) of the Securities Exchange Act of 1934 during the preceding 12 months (or for such shorter period that the Registrant was required to file such reports), and (2) has been subject to such filing requirements for the past 90 days.
Indicate by check mark whether the Registrant has submitted electronically every Interactive Data File required to be submitted pursuant to Rule 405 of Regulation S-T (§232.405 of this chapter) during the preceding 12 months (or for such shorter period that the Registrant was required to submit such files).
Indicate by check mark whether the registrant is a large accelerated filer, an accelerated filer, a non-accelerated filer, smaller reporting company, or an emerging growth company. See the definitions of “large accelerated filer,” “accelerated filer,” “smaller reporting company,” and “emerging growth company” in Rule 12b-2 of the Exchange Act.
|
Large accelerated filer |
☐ |
Accelerated filer |
☐ |
|
|
☒ |
Smaller reporting company |
|
|
Emerging growth company |
|
|
|
If an emerging growth company, indicate by check mark if the registrant has elected not to use the extended transition period for complying with any new or revised financial accounting standards provided pursuant to Section 13(a) of the Exchange Act.
Indicate by check mark whether the registrant has filed a report on and attestation to its management’s assessment of the effectiveness of its internal control over financial reporting under Section 404(b) of the Sarbanes-Oxley Act (15 U.S.C. 7262(b)) by the registered public accounting firm that prepared or issued its audit report.
Indicate by check mark whether the Registrant is a shell company (as defined in Rule 12b-2 of the Exchange Act). Yes
The aggregate market value of the voting and non-voting common equity held by non-affiliates of the Registrant was approximately $
The number of shares of Registrant’s common stock outstanding as of March 11, 2022 was
DOCUMENTS INCORPORATED BY REFERENCE
Portions of the Registrant’s definitive Proxy Statement relating to its 2022 Annual Meeting of Stockholders (Proxy Statement) are incorporated by reference into Part III of this Annual Report on Form 10-K (Annual Report) where indicated. The Proxy Statement will be filed with the U.S. Securities and Exchange Commission within 120 days after the end of the fiscal year to which this Annual Report relates.
Table of Contents
|
|
|
Page |
|
PART I |
|
|
|
Item 1. |
5 |
|
|
Item 1A. |
80 |
|
|
Item 1B. |
138 |
|
|
Item 2. |
138 |
|
|
Item 3. |
138 |
|
|
Item 4. |
138 |
|
|
|
|
|
|
PART II |
|
|
|
Item 5. |
139 |
|
|
Item 6. |
140 |
|
|
Item 7. |
Management’s Discussion and Analysis of Financial Condition and Results of Operations |
141 |
|
Item 7A. |
154 |
|
|
Item 8. |
155 |
|
|
Item 9. |
Changes in and Disagreements With Accountants on Accounting and Financial Disclosure |
181 |
|
Item 9A. |
181 |
|
|
Item 9B. |
181 |
|
|
Item 9C. |
Disclosure Regarding Foreign Jurisdictions that Prevent Inspections |
181 |
|
|
|
|
|
PART III |
|
|
|
Item 10. |
182 |
|
|
Item 11. |
182 |
|
|
Item 12. |
Security Ownership of Certain Beneficial Owners and Management and Related Stockholder Matters |
182 |
|
Item 13. |
Certain Relationships and Related Transactions, and Director Independence |
182 |
|
Item 14. |
182 |
|
|
|
|
|
|
PART IV |
|
|
|
Item 15. |
183 |
|
|
Item 16 |
186 |
i
SPECIAL NOTE REGARDING FORWARD-LOOKING STATEMENTS
This Annual Report on Form 10-K (Annual Report) contains forward-looking statements that involve substantial risks and uncertainties. All statements other than statements of historical facts contained in this Annual Report could be deemed forward-looking statements, including those statements highlighted below. In some cases, you can identify these statements by forward-looking words such as “aim,” “anticipate,” “believe,” “continue,” “could,” “estimate,” “expect,” “intend,” “may,” “might,” “plan,” “potential,” “predict,” “should,” “would,” or “will,” the negative of these terms, and other comparable terminology. These forward-looking statements, which are subject to risks, include, but are not limited to, statements about:
|
|
• |
our expectations regarding the potential market size and size of the potential patient populations for our product candidates and any future product candidates, if approved for commercial use; |
|
|
• |
our clinical and regulatory development plans; |
|
|
• |
our expectations with regard to the results of our preclinical studies, future clinical trials, and research and development programs, including the timing and availability of data from such studies and trials; |
|
|
• |
the timing of commencement of future preclinical studies, clinical trials, and research and development programs; |
|
|
• |
our ability to acquire, discover, and develop product candidates and advance them into, and successfully complete, clinical trials; |
|
|
• |
our intentions with respect to and our ability to establish collaborations or partnerships; |
|
|
• |
the timing or likelihood of regulatory filings and approvals for our product candidates; |
|
|
• |
our commercialization, marketing, and manufacturing expectations, including with respect to the buildout of our manufacturing facility and capabilities and the timing thereof; |
|
|
• |
impact of future regulatory, judicial, and legislative changes or developments in the United States and foreign countries; |
|
|
• |
our intentions with respect to the commercialization of our product candidates; |
|
|
• |
the pricing and reimbursement of our product candidates, if approved; |
|
|
• |
the potential effects of public health crises, such as the ongoing COVID-19 pandemic, on our preclinical and clinical programs and business; |
|
|
• |
our expectations regarding the impact of the ongoing COVID-19 pandemic on our business; |
|
|
• |
the implementation of our business model and strategic plans for our business and product candidates, including additional indications which we may pursue; |
|
|
• |
our ability to effectively manage our growth, including our ability to retain and recruit personnel, and maintain our culture; |
|
|
• |
the scope of protection we are able to establish and maintain for intellectual property rights covering our product candidates, including the projected terms of patent protection; |
|
|
• |
estimates of our expenses, future revenue, capital requirements, needs for additional financing, and ability to obtain additional capital; |
|
|
• |
our expected use of proceeds from our initial public offering and our existing cash, cash equivalents, and marketable securities; |
|
|
• |
the performance of our third-party suppliers and manufacturers; |
|
|
• |
our future financial performance; |
|
|
• |
our expectations regarding the time during which we will be an emerging growth company under the JOBS Act; and |
|
|
• |
developments and projections relating to our competitors and our industry, including competing products. |
We have based these forward-looking statements largely on our current expectations, estimates, forecasts, and projections about future events and financial trends that we believe may affect our financial condition, results of operations, business strategy, and financial needs. In light of the significant uncertainties in these forward-looking statements, you should not rely upon forward-looking statements as predictions of future events. Although we believe that we have a reasonable basis for each forward-looking statement contained in this Annual Report, we cannot guarantee that the future results, levels of activity, performance, or events and circumstances reflected in the forward-looking statements will be achieved or occur at all. You should refer to the sections titled “Risk Factors” and “Management’s Discussion and Analysis of Financial Condition and Results of Operations” for a discussion
1
of important factors that may cause our actual results to differ materially from those expressed or implied by our forward-looking statements. Other sections of this Annual Report may include additional factors that could harm our business and financial performance. New risk factors emerge from time to time, and it is not possible for our management to predict all risk factors, nor can we assess the impact of all factors on our business or the extent to which any factor, or combination of factors, may cause actual results to differ materially from those contained in, or implied by, any forward-looking statements. Except as required by law, we undertake no obligation to publicly update any forward-looking statements, whether as a result of new information, future events, or otherwise.
2
RISK FACTOR SUMMARY
Investing in our securities involves a high degree of risk. Below is a summary of material factors that make an investment in our securities speculative or risky. Importantly, this summary does not address all of the risks that we face. Additional discussion of the risks summarized in this Risk Factor Summary, as well as other risks that we face, can be found under the heading “Risk Factors” in Part I of this Annual Report.
Our business is subject to a number of risks of which you should be aware before making a decision to invest in our common stock. These risks include, among others, the following:
|
|
• |
Our ex vivo and in vivo cell engineering platforms are based on novel technologies that are unproven and may not result in approvable or marketable products. This uncertainty exposes us to unforeseen risks, makes it difficult for us to predict the time that will be required for the development and potential regulatory approval of our product candidates, and increases the risk that we may ultimately not be successful in our efforts to use and expand our technology platforms to build a pipeline of product candidates. |
|
|
• |
If we are unable to successfully identify, develop, and commercialize any product candidates, or experience significant delays in doing so, our business, financial condition, and results of operations will be materially adversely affected. |
|
|
• |
While we believe our pipeline will yield multiple investigational new drug applications (INDs), we may not be able to submit INDs to commence clinical trials on the timelines we expect, and even if we are able to submit an IND, the United States Food and Drug Administration (FDA) may not permit us to proceed with clinical trials. |
|
|
• |
We may not realize the benefits of technologies that we have acquired, or will acquire in the future, or any collaborative or licensing arrangement or other strategic transactions that we have or will consummate. If we fail to enter into new strategic relationships, our business, financial condition, commercialization prospects, and results of operations may be materially adversely affected. |
|
|
• |
Our ability to develop our cell engineering platforms and product candidates and our future growth depends on retaining our key personnel and recruiting additional qualified personnel. |
|
|
• |
We may encounter difficulties in managing our growth as we continue to expand our development and regulatory capabilities, which could disrupt our operations. |
|
|
• |
The use of human stem cells exposes us to a number of risks in the development of our human stem cell-derived products, including an inability to obtain suitable donor material from eligible and qualified human donors, restrictions on the use of human stem cells, as well as ethical, legal, and social implications of research on the use of stem cells, any of which could prevent us from completing the development of or commercializing and gaining acceptance for our products derived from human stem cells. |
|
|
• |
All of our product candidates are in preclinical development and none have commenced clinical development. Preclinical and clinical drug development is a lengthy and expensive process with uncertain timelines and uncertain outcomes. If preclinical studies or clinical trials of any of our product candidates are prolonged or delayed, we may be unable to obtain required regulatory approvals and commercialize such product candidates on a timely basis or at all. |
|
|
• |
Our future clinical trials may fail to demonstrate substantial evidence of the safety and efficacy of our product candidates, including any future product candidates, which would prevent, delay or limit the scope of regulatory approval and commercialization of such product candidates. |
|
|
• |
Our product candidates may have serious adverse, undesirable, or unacceptable side effects or other properties that may delay or prevent marketing approval. If a product candidate receives regulatory approval, and such side effects are identified following such approval, the commercial profile of any approved label may be limited, or we may be subject to other significant negative consequences following such approval. |
|
|
• |
The manufacture of our product candidates is complex. We or our third-party CDMOs may encounter difficulties in production, which could delay or entirely halt our or their ability to supply our product candidates for clinical trials or, if approved, for commercial sale. |
|
|
• |
We are exposed to a number of risks related to the supply chain for the materials required to manufacture our product candidates. |
|
|
• |
We rely on, and expect to continue to rely on, third parties to perform certain activities, including research and preclinical studies, manufacture of our product candidates and materials used to manufacture our product candidates, and the conduct of various aspects of our planned clinical trials. Any failure of such third parties to perform their obligations to us, including in accordance with our timelines or applicable regulatory requirements, could materially harm our business. |
3
|
|
• |
Our success depends on our ability to protect our intellectual property rights and our proprietary technologies. |
|
|
• |
We depend on intellectual property licensed from third parties. If we breach our obligations under these agreements or if any of these agreements is terminated, we may be required to pay damages, lose our rights to such intellectual property and technology, or both, which would harm our business. |
|
|
• |
Our internal computer systems, or those used by our third-party research institution collaborators, CROs, CDMOs, or other contractors or consultants, may fail or suffer security breaches. |
|
|
• |
The development and commercialization of biopharmaceutical products is subject to extensive regulation, and the regulatory approval processes of the FDA and comparable foreign authorities are lengthy, time-consuming, and inherently unpredictable. If we are unable to obtain regulatory approval for our product candidates on a timely basis, or at all, our business will be substantially harmed. |
|
|
• |
We are a preclinical-stage biotechnology company and have incurred significant losses since our inception, and we expect to incur losses for the foreseeable future. We have no products approved for commercial sale and may never achieve or maintain profitability. |
|
|
• |
We will require additional funding in order to finance our operations. If we are unable to raise capital when needed, or on acceptable terms, we could be forced to delay, reduce, or eliminate our product development programs or commercialization efforts. |
|
|
• |
Our success payment and contingent consideration obligations may result in dilution to our stockholders, drain our cash resources, or cause us to incur debt to satisfy the payment obligations. |
|
|
• |
Our limited operating history may make it difficult to evaluate our prospects and likelihood of success. |
|
|
• |
We or the third parties upon whom we depend may be adversely affected by natural disasters, public health epidemics, such as the ongoing COVID-19 pandemic, telecommunications or electrical failures, geo-political actions, including war and terrorism, political and economic instability, and other events beyond our control, and our business continuity and disaster recovery plans may not adequately protect us from a serious disaster. |
4
PART I
Item 1. Business.
Overview
We were founded on the belief that engineered cells will be one of the most important transformations in medicine over the next several decades. The burden of diseases that can be addressed at their root cause through engineered cells is significant. We view engineered cells as having the potential to be as therapeutically disruptive as biologics to clinical practice. Our long-term aspirations are to be able to control or modify any gene in the body, to replace any cell that is damaged or missing, and to markedly improve access to cellular and gene-based medicines. We have brought together an experienced group of scientists, engineers, and company builders and combined them with the necessary technologies to move this vision forward. We are developing ex vivo and in vivo cell engineering platforms to revolutionize treatment across a broad array of therapeutic areas with unmet treatment needs, including oncology, diabetes, central nervous system (CNS) disorders, cardiovascular diseases, and genetic disorders, among others. Our platform progress, broad capabilities, and strong balance sheet enable us to execute on a broad vision, with a goal of submitting our first INDs in 2022, with the opportunity to submit multiple INDs per year beyond 2022.
We believe the time is right to develop engineered cell therapies across a broad range of therapeutic areas. The field has seen initial clinical proof of concept for gene and cell replacement approaches across multiple diseases, including cancer and certain genetic disorders, through the application of adeno-associated virus (AAV) based gene therapies, autologous CAR T cell therapies, and autologous and allogeneic grafts/transplants. While such existing approaches have limitations, they provide evidence that a broad range of ex vivo and in vivo engineered cells can have transformative clinical potential in at least a subset of patients. Substantial progress in the understanding of genetics, gene editing, gene control, protein engineering, stem cell biology, immunology, process analytics, and computational biology have converged to create an opportunity to markedly increase the breadth and depth of the potential impact of genetic and cellular medicines.
We are seeking to overcome these existing limitations of gene and cell therapy through our ex vivo and in vivo cell engineering platforms, both of which may facilitate the development of therapies that can transform the lives of patients by repairing cells in the body when possible and replacing them when needed. For ex vivo therapies, where diseased cells are damaged or missing entirely and an effective therapy needs to replace the entire cell, a successful therapeutic requires large-scale manufacturing of cells that engraft, function, and persist in the body. Of these, we view persistence as the greatest limitation to dramatically expanding the impact of this class of therapeutics. We believe that product candidates developed with our ex vivo cell engineering platform, which utilizes hypoimmune allogeneic cells that can “hide” from the patient’s immune system, can address this fundamental limitation and unlock a wave of disruptive therapeutics. For in vivo therapies, where the desire is to repair and control genes in the body, a successful product candidate requires both gene modification and in vivo delivery of the therapeutic payload. Of these, we view effective in vivo delivery as the greatest limitation to dramatically expanding the impact of this class of therapeutics. To this end, our initial focus is on cell-specific delivery as well as increasing the diversity and size of payloads.
We believe we have the potential to develop transformative engineered cells as medicines because of our people and our capabilities:
Our people are the most important strength of the company. We have assembled a diverse group of experienced company builders, scientists, manufacturing scientists, engineers, and operators to execute our business plan.
|
|
• |
Experienced Company Builders. We have numerous individuals with vast experience in building disruptive biotech companies. Our Founder and Chief Executive Officer, Dr. Steve Harr, was previously CFO of Juno Therapeutics, helping to build the company and its CAR T cell therapy platform until its acquisition. He is a physician-scientist with experience in basic research, clinical medicine, finance, company building, and operations. Our Chairman of the Board and co-founder, Hans Bishop, is an experienced company builder and operator with success across a number of companies. Our executive team is composed of multiple individuals with deep experience building high growth, disruptive companies, including Christian Hordo, Chief Business Officer, who previously ran Business Development and the Myeloma program at Juno Therapeutics, and Robin Andrulevich, Chief People Officer, who has held key senior leadership roles at Amazon, Google, and Juno Therapeutics. |
|
|
• |
Leading Scientists. We believe that in order to be successful in drug development for engineered cells, significant investments in infrastructure and cross-functional capabilities need to be coupled with deep scientific expertise in the cell types of interest within each program. Our leadership team includes multiple world-class scientists, including researchers who have made seminal discoveries in gene delivery, immunology, CAR T cells, gene editing, and stem cell biology. These include Drs. Richard Mulligan, Terry Fry, Ed Rebar, Chuck Murry, Sonja Schrepfer, Steve Goldman, and Jagesh Shah. We have surrounded this team of discovery scientists with drug developers experienced in advancing product candidates through the development process with expertise in areas such as pharmacology, toxicology, regulatory, clinical |
5
|
|
development, and clinical operations. These include Drs. Sunil Agarwal, Donna Dambach, Ke Liu, Paul Brunetta, and Ms. Farah Anwar. |
|
|
• |
Experienced Manufacturing Scientists, Engineers, and Operators. Since our founding, we have proactively assembled manufacturing sciences and operations expertise on our board, on our executive team, and across the company. Our manufacturing organization is led by Dr. Stacey Ma, an experienced executive with over two decades of manufacturing leadership, contributing to the commercialization of over ten products across multiple modalities. |
|
|
• |
Board and Investors with Shared Long-Term Vision. Our board of directors is composed of renowned company builders, scientists, drug developers, and investors who share our long-term vision of advancing engineered cells as medicine to change the lives of patients. This has enabled our strategy of consolidating technologies, assets, and people to expand the potential impact of our long-term vision. |
Our capabilities enable us to take a comprehensive approach to the most important and difficult aspects of engineering cells. We are pursuing ex vivo and in vivo cell engineering and can leverage the synergistic proficiencies required to succeed in both approaches. We believe we can capitalize on the shared expertise and infrastructure between the platforms to maximize the potential success and the reach of our transformative therapies. We have built deep internal capabilities across a wide range of areas focused on solving the most critical limitations in engineering cells including:
|
|
• |
Gene Delivery. We believe our delivery technologies have broad potential, with both near-term and long-term applications across a number of indications. We are investing in technologies that allow payload delivery to specific cell types, increase the diversity and size of payloads, enable repeat dosing of patients, and increase the volume of distribution inside the body in order to target and access more diverse cells. |
|
|
• |
Gene Modification. The ability to knock-out, knock-in, modify, and control expression of genes is fundamental to our platforms’ success. We have hired world-class scientists with experience in all of these capabilities and across multiple modalities. We are building internal capabilities that enable high throughput cell engineering and gene editing and control using multiple technologies through use of natural systems, protein engineering, and synthetic biology. We believe our capabilities across multiple modalities will allow us to utilize the appropriate system for the biologic problem of interest. We are developing proprietary gene editing capabilities as well as seeking strategic partnerships in key areas. |
|
|
• |
Immunology. The immune system can be harnessed to treat multiple diseases, and it can also limit the therapeutic effect of most cell- and gene-based therapies. Understanding and harnessing the immune system can have a broad impact across our ex vivo and in vivo cell engineering portfolio. We are investing in our people and technologies to harness the immune system, particularly T cells, for the treatment of cancer and other diseases. Additionally, our hypoimmune technology has the potential to hide cells from the immune system, unlocking the potential of allogeneic ex vivo therapies for the treatment of numerous diseases. |
|
|
• |
Stem Cell and Disease Biology. Developing our platforms into therapies for patients requires a deep understanding of both cell and disease biology. Furthermore, we are investing significantly in our people and the technologies that enable the differentiation of pluripotent stem cells into mature cells that can be used as therapeutics. In each therapeutic area we intend to pursue, we have brought in-house senior world-class scientists to lead our efforts, and our research teams have significant experience in various areas of biology. |
Our ex vivo and in vivo Cell Engineering Platforms
The advent of recombinant DNA technology in the 1970s ushered in a new era of therapeutics, enabling the synthetic manufacture of human protein therapies at scale for the first time. However, the critical inflection point occurred when key technological advancements eventually enabled the broad development of monoclonal antibodies with suitable therapeutic properties. These advancements, combined with progress in understanding disease biology, allowed biologics to become the second largest therapeutic class. We believe engineered cells are at a similar inflection point, with key recent technological advancements providing the potential for the broad applicability of this therapeutic class.
Ex vivo cell engineering
Engineering cells ex vivo requires the ability to engineer and manufacture cells at scale and then deliver them to the patient, so that they engraft, function appropriately, and have the necessary persistence in the body. Our goal for ex vivo cell engineering is to replace any cell in the body with cells that engraft, function, and persist over time, and to manufacture those cells cost-effectively at scale. Our ex vivo cell engineering platform utilizes our hypoimmune technology to create cells that can “hide” from the patient’s immune system to enable persistence of allogeneic cells. We are striving to make therapies utilizing pluripotent stem cells with our hypoimmune genetic modifications as the starting material, which we then differentiate into a specific cell type, such as a pancreatic beta cell, before treating the patient. Additionally, for cell types for which effective differentiation protocols from a stem cell have not yet been developed, such as T cells, instead of starting from a pluripotent stem cell, we can utilize a fully differentiated allogeneic cell,
6
sourced from a donor, as the starting material to which we then apply our hypoimmune genetic modifications. Our goal is to manufacture genetically modified cells that are capable of both replacing the missing cell and evading the patient’s immune system. We are now applying our technologies to make cell products for the treatment of multiple diseases.
In vivo cell engineering
Engineering cells in vivo requires the development of both an appropriate delivery vector as well as a payload to effectively modify the cell. Our goal for in vivo cell engineering is to repair and control the genes of any cell in the body. The ultimate aim is to achieve the delivery of any payload, to any cell, in a specific and repeatable way. Our in vivo cell engineering platform harnesses fusogen technology, which targets cell surface receptors, and thereby can enable cell specific delivery for a meaningful number of different cell types. Using our fusogen technology, we have shown in preclinical studies that we can specifically target numerous cell surface receptors that, when combined with delivery vehicles to form fusosomes, allow cell-specific delivery across multiple different cell types.
Our Portfolio Strategy
We believe the potential applications of our platforms are vast. To prioritize programs for our ex vivo and in vivo engineering pipeline we have used the following strategies:
|
|
• |
minimize biology risk where there is platform risk, or in other words, prioritize opportunities where success with our platform should lead to success in addressing the underlying disease; |
|
|
• |
prioritize program investments in diseases where the strengths of our ex vivo and in vivo cell engineering platforms can address the key limitations of existing therapeutic approaches; |
|
|
• |
focus on conditions of high unmet need, including the most grievous diseases; and |
|
|
• |
prioritize efforts where success in one area begets success in others. |
Our Pipeline
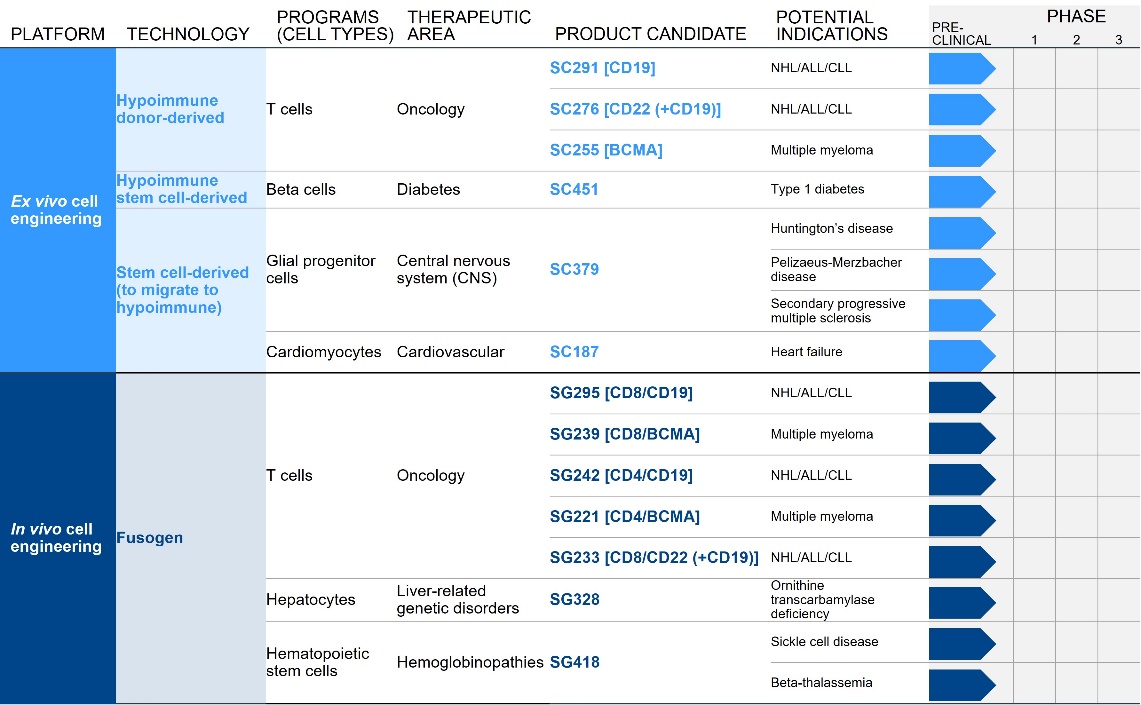
We are developing a broad pipeline of product candidates focused on creating transformative ex vivo and in vivo engineered cell therapies across a range of therapeutic areas. We are in the early stages of development across a broad pipeline of product candidates, all of which are currently in the preclinical stage of development and are summarized below:
Each of our initial programs provides the potential for meaningful standalone value while also supporting our potential ability to further exploit our platforms into broadly applicable medicines.
Our most advanced hypoimmune product candidate is SC291, a CD19-directed allogeneic CAR T program for NHL, CLL, and ALL. This program is designed to address the major limitation of existing allogenic CAR T cell therapies: evasion of host versus
7
graft responses (HvGR), which occurs when a patient’s immune system kills the transplanted T cells, limiting the potential benefit of the therapy. One approach to avoid HvGR has been to effectively eliminate a patient’s immune system for a short period using chemotherapy, which puts the patient at risk for severe infections. Further, the patient’s immune system will inevitably recover, which will lead to the immune system eliminating the CAR T cells, limiting the effectiveness of the therapy. Our hypoimmune technology is designed to hide cells from the patient’s immune system, giving our allogeneic CAR T cell program the potential to create medicines that persist longer in patients and avoid the risks associated with higher doses of chemotherapy. Our goal is to submit an IND for SC291 in 2022, and initial clinical success would unlock meaningful standalone value in the development of SC291 in NHL/CLL/ALL which has the potential to address scalability challenges of autologous therapies. Additionally, this initial clinical success would support and validate the expansion of our allogenic CAR T efforts, which include a CD22-targeting allogeneic CAR T which could be combined with targeting CD19 (SC276) and offers the potential benefit of higher and more durable complete response rates, and a BCMA directed CAR T (SC255) in multiple myeloma. Furthermore, initial clinical success would also support the validation of the hypoimmune platform overall, which is being actively deployed internally across a number of other therapeutic areas beyond oncology.
Our next most advanced hypoimmune product candidate is SC451, PSC-derived pancreatic beta cells for the treatment of diabetes, with an initial focus on Type I diabetes mellitus (T1DM). Almost 1.6 million people in the United States, and 2.4 million in Europe have T1DM. T1DM is a disease in which a patient’s immune system attacks and kills pancreatic beta cells, leading to complete loss of insulin production in affected individuals. Patients need to take multiple insulin injections every day for life, and, while insulin has a profoundly positive impact on patients, people with T1DM have approximately 15 years shorter life expectancies than people without diabetes and are consistently at risk for complications such as coma, stroke, myocardial infarction, kidney failure, and blindness from poorly controlled blood glucose. We and our collaborators have shown that we can develop high quality beta cells that, when transplanted, normalize blood glucose and cure diabetes in animal models. We have also shown that our hypoimmune cells induce no systemic immune response, even in NHPs with a pre-existing immune response to non-hypoimmune cells. As a result, we believe our stem cell derived hypoimmune pancreatic cells have the potential to create a disruptive treatment for T1DM, offering patients life-long normal blood glucose without immunosuppression. We are working through the process development and IND-enabling studies to allow for an IND submission for SC451 as early as 2023.
Our most advanced fusosome product candidate is SG295, which targets CD19+ cancer cells, including NHL, CLL, and ALL. This program provides us with an opportunity to develop potential product candidates to expand access to CAR T cell therapy to many more patients in need. In addition, we believe the ability to deliver a payload encoding a CAR to a T cell without meaningful ex vivo manipulation has the potential to improve effectiveness over ex vivo manufactured CAR T cell products. These approaches should result in the generation of therapeutically active CAR T cells without the complexities and delays associated with the process of T cell collection and ex vivo manufacturing. Furthermore, the ex vivo expansion in the presence of high cytokine concentrations, while necessary for the manufacture of approved CAR T cell products, also contributes to marked changes in T cell quality that may not be therapeutically beneficial. The generation of a CAR T cell within the natural physiological environment in vivo has the potential to improve the quality of the CAR T cell generated, potentially improving both efficacy and the side effect profile. Finally, the effectiveness of ex vivo manufactured CAR T cells is dependent on the administration of a lymphodepleting preparative regimen prior to infusion to facilitate expansion of the CAR T cell product, which can have meaningful adverse safety implications. We do not expect to need a lymphodepleting regimen prior to in vivo delivery of the CAR gene, as our goal is to expose our fusosomes to as many T cells in the body as possible. Our goal is to submit an IND for SG295 in 2022 and initial clinical success would unlock meaningful standalone value in the development of SG295 in NHL/CLL/ALL Additionally, this initial clinical success would support and validate the expansion of our in vivo CAR T efforts and support the validation of our fusosome platform overall, which is being actively deployed internally across a number of other therapeutic areas beyond oncology with the goal of targeted delivery of DNA and gene editing machinery to specific cells in vivo.
Our ex vivo Cell Engineering Platform
Overview
Ex vivo cell engineering aims to treat human disease by engrafting new cells to replace diseased cells that are damaged or missing in patients. Historically there have been four key challenges to ex vivo cell engineering:
|
|
• |
engraftment of the right cell in the right environment; |
|
|
• |
appropriate function of the cells, necessitating an understanding of and ability to produce the desired cell phenotype; |
|
|
• |
persistence of the cells in the host, particularly by overcoming immune rejection; and |
|
|
• |
manufacturing the desired cell in the quantities required. |
8
Our ex vivo cell engineering platform seeks to address these four challenges and is focused on engineering hypoimmune cells that engraft, function, and persist in patients by evading immune rejection. These are derived from cell sources that are scalable and we believe that continued progress with this platform has the potential to create broad access for patients.
Our Approach to Building our ex vivo Cell Engineering Platform
We have approached the development of our ex vivo cell engineering platform by investing in solutions to address the key challenges outlined above:
|
|
• |
Stem cell and disease biology. We believe that it is critical to have expertise in the developmental biology of stem cell differentiation and a deep understanding of the desired cell phenotype biology of stem cell differentiation in order to generate cells that function appropriately, as well as a deep understanding of the desired cell phenotype. The latter requires expertise in normal and disease biology. Furthermore, clinical understanding of disease pathology and transplant medicine is required to determine how to engraft the right cell in the right environment. Each of our programs is led by a prominent clinician-scientist with deep expertise in both cell therapy and disease biology, including Dr. Terry Fry, our Senior Vice President, Head of T Cell Therapeutics, for T cells, Dr. Steve Goldman, our Senior Vice President, Head of CNS Therapy, for glial cells, and Dr. Chuck Murry, our Senior Vice President, Head of Cardiometabolic Cell Therapy, for cardiomyocytes and beta cells. |
|
|
• |
Immunology and gene modification. We believe that a deep understanding of the immunological response to engineered cells is essential to unlocking the potential of ex vivo therapies. This effort is led by Dr. Sonja Schrepfer, our Senior Vice President, Head of Hypoimmune Platform, and draws from decades of research. We have licensed technologies from University of California San Francisco, Harvard University, Washington University, and others to enable this effort. In addition, in order to create successful hypoimmune cells, we are investing in building out our gene editing, modification, and insertion capabilities, led by Dr. Ed Rebar, our Senior Vice President, Chief Technology Officer. |
|
|
• |
Manufacturing. We are investing proactively in process development, including process optimization and scale up, analytical development, CMC regulatory, supply chain, quality, and other manufacturing sciences in order to develop processes that can enable scalable manufacturing of cell therapies and broad patient access. We have also built a pilot manufacturing plant in South San Francisco, California and entered into a long-term lease agreement for a facility in Fremont, California, where we intend to build our own clinical trial and commercial Good Manufacturing Practice (GMP) manufacturing capabilities. We are also investing to access high quality donor-derived T cells and GMP-grade pluripotent stem cell lines for our programs. These manufacturing efforts are led by Dr. Stacey Ma, our Executive Vice President, Technical Operations. |
Our Approach to Building our ex vivo Cell Engineering Portfolio
We have prioritized cell types for our programs where:
|
|
• |
high unmet need can be addressed by cell replacement; |
|
|
• |
existing proof of concept in humans and/or animal models demonstrates that cell transplantation should have a clinical benefit; |
|
|
• |
evidence exists that the cell type can be successfully differentiated from pluripotent stem cells and that such stem cell-derived cells can function appropriately in vivo; |
|
|
• |
there has been the ability to hire or partner with one of the world experts in the field to ensure our programs are rooted in a deep understanding of the underlying cell and disease biology; and |
|
|
• |
evading immune system rejection via the hypoimmune technology is either not required initially, but would be disruptive over time (such as cardiomyocytes) or is the critical missing element to developing a cell therapy (such as beta cells). |
Based on this prioritization, we are initially focused on four cell types: T Cells, Beta cells, GPCs, and Cardiomyocytes.
Historical context of ex vivo therapy
Blood transfusions have been a standard treatment for many patients for over 100 years. The first successful kidney transplant occurred in 1954, followed by the first successful heart transplant in 1967, demonstrating the transformative clinical potential of replacing damaged or missing cells in the body. Surgical enhancements have improved the success of engraftment, but lack of organ access, a complex surgical procedure, and immune rejection of the donated organs have limited the impact of these procedures.
9
Progress in immunosuppressive regimens, such as the development of cyclosporine, has improved organ survival rates. However, substantial side effects and the fact that many patients are ineligible or non-compliant have reduced their impact.
Ultimately, the field has looked for a scalable source of therapeutic cells that can be accessed broadly at a manageable cost, as well as cells that can evade immune rejection without immunosuppression. The advent of stem cell technology and subsequent improvements in methods to generate functional differentiated cells at scale have the potential to address the shortage of donor tissues and organs. In addition, over the past decade a deeper understanding of the immunology of host versus graft responses, coupled with novel techniques to manipulate the immunological profile of cells via gene editing, have raised the prospect that ex vivo engineered cells can significantly benefit patients without the requirement for significant immunosuppression.
Sources of allogeneic cells
There are three main potential sources of allogeneic cells, or cells that do not originate from the patient, and therefore have the potential to be manufactured and supplied at scale. These are embryonic stem cells (ESCs), iPSCs, and donor-derived cells. Our portfolio currently reflects a mix of sources, with the ambition of transitioning primarily to iPSCs over time.
Embryonic Stem Cells
The recognition that every cell in the body originates from a zygote, or fertilized egg, led to the research and ultimate discovery of human ESCs, with the derivation of the first human ESC line in 1998. ESCs are pluripotent stem cells which can potentially differentiate into any cell type and are derived from the inner cell mass of a blastocyst or pre-implantation stage embryo. They are typically cultured in vitro and grown through cycles of cell division, known as passages, until a line of cells is established that can proliferate without differentiating, and retain their pluripotency while remaining well characterized, including free from potentially deleterious genetic mutations. Because pluripotent stem cells can divide indefinitely without exhaustion, an ESC line can be used to generate cell banks, consisting of large numbers of well-characterized vials of cells, that can be frozen and stored for future use.
Induced Pluripotent Stem Cells
The discovery that mature, differentiated cells can be reprogrammed to be the equivalent of an ESC and capable of generating any cell type in the body, has led to the research and ultimate development of human iPSCs, providing an alternative option as a source of stem cells for use in ex vivo engineered cells. A key scientific step was the breakthrough in 2006 demonstrating that mature cells could be reprogrammed via the expression of a small number of genes to result in pluripotent cells These iPSCs have similar potential to ESCs to be used as an indefinitely renewable cell bank for manufacturing of cell-based therapies.
Donor-Derived Allogeneic Cells
Another source of cells, which we utilize in our T cell program, comes from mature donor-derived allogeneic cells. While these cells are neither pluripotent nor from an infinitely renewable source, T cells can be obtained as mature cells from human donors at scale. The use of donor-derived cells for our T cell program should allow us to rapidly advance the program towards the clinic with the implementation of our hypoimmune technology.
Approach to Sources of Allogeneic Cells
We are focused on iPSCs as the starting material for our programs, which offers regulatory and cultural advantages to ESCs, and scale and product consistency advantages to donor-derived allogeneic cells. Our portfolio currently reflects a mix of sources, which is primarily driven by historical factors as well as current better characterization of genomic stability through differentiation. Our ambition is to transition primarily to iPSCs over time.
Crucial aspects of developing allogeneic cells from any source include the thorough characterization of the cells, a comprehensive understanding of the global regulatory environment, and an ability to maintain cells under the required conditions, such as current GMP (cGMP), at various stages of the manufacturing processes. We believe our early investment in building capabilities in the science and manufacturing of these cells will increase our likelihood of success. This investment is anticipated to
10
yield sources of cells suitable for the global clinical development and commercialization of ex vivo engineered cells for a broad patient population, in line with our vision to democratize access.
Background on Immunological Barriers to ex vivo Therapies and Current Limitations
Starting with studies in renal transplantation in the early 1900s, it became clear that there were immunological factors preventing successful transplantation. Initially, it was suspected to be mediated by an antibody response, but in the 1950s it was discovered that cell-mediated immune pathways also play a critical role in transplant rejection.
Further studies established T cells as playing a key role in the host immune response to transplant. T cells belong to the “adaptive” immune system, recognizing and eliminating “non-self” cells via recognition of differences in cell-surface proteins encoded by the major histocompatibility (MHC) locus. There are two types of MHC molecules: MHC class I, expressed on the surface of almost all nucleated cells, and MHC class II, expressed constitutively on professional antigen presenting cells (APC), including macrophages and dendritic cells. Expression of MHC class II is also induced in many additional cells in the context of inflammation. MHC class I molecules typically display peptides on the cell surface from degraded intracellular proteins. Cells display peptides from normal “self” proteins on MHC class I, which typically will not activate an immune response due to a process called tolerance, where the body recognizes these peptides as “self”. However, if a cell displays a peptide from a foreign or mutated protein on MHC class I, for example as a result of a protein mutation, it may result in the activation of a cytotoxic T cell response specific to the peptide-MHC complex via the T cell receptor (TCR) on the T cell surface. The activated T cell then eliminates the cell. MHC class II molecules typically display peptides derived from phagocytosis of extracellular proteins on the surface of APCs. These peptide-MHC complexes interact with TCRs on helper T cells, such as CD4+ T cells, resulting in a downstream cellular and humoral immune response. The humoral immune response leads to antibody production against foreign proteins. In allogeneic transplants, the cellular and humoral processes can recognize proteins from the donor as “foreign”, resulting in an immune response to the transplant including potential elimination of the transplanted cells. In the allogeneic setting, MHC proteins can be highly immunogenic due to their inherent polymorphism, increasing the risk of the recognition of transplants as “foreign”. This underlies the basis for MHC typing and matching to assess and reduce the risk of organ transplant rejection.
Many groups have attempted to engineer cells that can evade the adaptive immune system, typically by downregulating or eliminating expression of MHC molecules on the surface of cells. While this can reduce the adaptive immune response to donor cells, the human immune system has evolved so that parts of the innate immune system will recognize cells missing MHC molecules and eliminate them. For example, natural killer (NK) cells express receptors known as inhibitory killer-cell immunoglobulin-like receptors (inhibitory KIRs). KIRs recognize self MHC class I molecules on the surface of cells and provide inhibitory signals to the NK cells to prevent their activation. Cells missing MHC class I molecules are correspondingly eliminated by NK cells because of the lack of inhibitory KIR signaling and a resulting cytolytic activation. Known as the “missing self-hypothesis,” this important redundancy in immunology enables the elimination of virally infected or transformed cells that have downregulated MHC class I, but also has complicated the development of allogeneic cells as broadly applicable therapeutics. Our hypoimmune technology seeks to engineer cells to avoid immune rejection by addressing both the adaptive and innate immune response.
There are three key strategies that have been utilized to date to overcome immune rejection, with limited success:
|
|
• |
Immune Suppression. Cyclosporine and other molecules that suppress T cell responses are commonly used, and many patients have been helped by the approaches in areas such as an organ transplantation. However, immune suppression often leads to significant systemic side effects, including a decreased ability to fight-off infections, increased susceptibility to cancer, and a wide variety of organ toxicities. Furthermore, patients typically require these on a lifelong basis, and any disruption in immunosuppression can rapidly trigger rejection. |
|
|
• |
Matching HLA Type. A second approach to overcoming immune rejection is to find a donor with a matched HLA type. HLA stands for human leukocyte antigen which, in humans, is a synonym for MHC. This approach addresses the root of the mechanism that the immune system uses to identify “non-self” cells and has achieved some success. Finding a matched donor, however, can be difficult and is usually limited to close relatives who are willing and able to donate. While some have advocated for creating large banks of cells that match a wide variety of HLA types, even with fully matched HLA class I and class II donors and recipients, there is a need for at least some immune suppression due to the presence of numerous minor antigen mismatches. |
|
|
• |
Autologous Approaches. More recently, researchers have pursued autologous approaches, where a patient’s own cells are modified and introduced back as a graft. These cells may avoid immune rejection as they would be recognized as “self.” Autologous approaches have demonstrated effectiveness in certain diseases, such as autologous CAR Ts for hematological malignancies, but these are limited in their adoption due to manufacturing cost and complexity. Furthermore, autologous approaches are generally limited to cells that exist in the patient in suspension, such as blood cells, and they cannot be applied to treat acute illnesses, such as myocardial infarction or stroke, due to the time it takes to prepare these cells for administration. |
11
Our Solution – Hypoimmune Technology
To address the challenge of immune rejection with allogeneic cell transplantation, we are developing our hypoimmune technology, utilizing gene modification to introduce permanent changes to the cells. We are applying the hypoimmune technology to both iPSCs, which can then be differentiated into multiple cell types, and to donor-derived allogeneic T cells, which has the goal of making potent CAR T cells at scale. Our goal with this technology is to transplant allogeneic cells into patients without the need for systemic immune suppression. We believe that enabling this capability has the potential to enable ex vivo engineered cells to become an important therapeutic modality alongside small molecules, protein biologics, and in vivo engineered cells.
Some of our scientific founders, including Dr. Sonja Schrepfer, our Senior Vice President, Head of Hypoimmune Platform, and their collaborators have worked on creating hypoimmune cells for well over a decade. A key insight was focusing on the phenomenon of fetomaternal tolerance during pregnancy. The fetus, despite having half its genetic material from the father, is not rejected by the mother’s immune system. However, after birth, few if any children would qualify as a matched donor for a cell or organ transplant for their mother. These scientists categorized the differences of the maternal-fetal border and systematically tested them to understand which, if any, of these were most important to immune evasion. They have tested these changes in both in vitro and in vivo animal models.
Designing Hypoimmune Cells
Our goal is to create a universal cell that is able to evade immune detection, regardless of cell type or transplant location. Our first-generation technology, which is progressing through late-stage animal confirmatory studies, combines the three gene modifications below to hide these cells from the host immune system:
|
|
• |
disruption of MHC class I expression; |
|
|
• |
disruption of MHC class II expression; and |
|
|
• |
overexpression of CD47, a protein that hides cells from the innate immune system, including macrophages and NK cells. |
Preclinical Development of Hypoimmune Cells
We and our licensors have carried out a series of experiments in various model systems of increasing immunological complexity. These included (i) transplanting undifferentiated mouse hypoimmune iPSCs- into MHC mismatched allogeneic mice, (ii) transplanting mouse hypoimmune iPSC-derived differentiated cells, such as endothelial cells, into MHC mismatched allogeneic mice, (iii) transplanting human hypoimmune iPSCs into MHC mismatched humanized allogeneic mice; and (iv) transplanting human hypoimmune iPSCs into non-human primates (NHPs). We are currently carrying out experiments transplanting NHP hypoimmune iPSC cells into NHPs as well as transplanting NHP hypoimmune iPSC-derived differentiated cells, such as cardiomyocytes, into allogeneic NHPs.
Each mouse experiment evaluated:
|
|
• |
whether hypoimmune cells can be successfully transplanted into the recipient without the need for immunosuppression and without eliciting an immune response; and |
|
|
• |
whether differentiated cells derived from our hypoimmune cells were successfully engrafted in the recipient without needing immunosuppression and without eliciting an immune response. |
We are investigating both human iPSCs in NHPs as well as NHP iPSCs in NHPs, as we want insights into how the NHP immune system reacts to each of these species. We have largely completed the study of human iPSCs and have early results from the NHP hypoimmune iPSC transplantation experiments. We are encouraged by data to date across species, with the NHP immune system most closely resembling the human immune system, representing the strictest test outside of testing these cells in humans. We are evaluating both iPSCs as well as differentiated cells transplanted into the microenvironment we intend to target in humans. Based on the results of these NHP studies, we expect to test these hypoimmune cells in humans as a next step.
Mouse iPSC-derived hypoimmune cells transplanted into MHC mismatched allogeneic mouse
Mouse hypoimmune iPSCs transplanted into an MHC mismatched allogeneic mouse were protected from the mouse immune system, and no evidence was seen of either adaptive or innate immune system activation. The control arm transplanted non-edited mouse iPSCs into MHC mismatched allogeneic mice, and, as expected, these non-edited mouse iPSCs were rapidly rejected by the recipient’s immune system with a robust adaptive immune response. In another experiment, the genes that code for MHC class I and MHC class II expression were knocked out. These modifications protected the cells from the recipient mouse’s adaptive immune
12
system, but NK cells rapidly killed the transplanted cells. These data highlight the importance of making all three gene modifications in order to protect cells from the immune system with an allogeneic transplant.
Next, to ensure that hypoimmune gene modifications protected differentiated cells and that these modifications did not impact the ability of iPSCs to differentiate into various cell types, commonly referred to as pluripotency, it was tested whether the hypoimmune iPSCs cells could be differentiate into three different cell types, function in vivo, and evade the host immune system. The three cell types were cardiomyocytes, endothelial cells, and smooth muscle cells. It was observed that hypoimmune iPSCs could successfully differentiate into all three cell types, the cells functioned in the mouse, and the transplanted cells survived for the full standard observation period with no evidence of immune system activations despite any immune suppression. Differentiated cells derived from non-edited iPSC cells led to immune activation in the host mouse, and they did not survive. These data provide initial proof of concept that iPSCs can be genetically modified, and differentiated into target cells that can engraft, function, and evade the recipient’s immune system following transportation.
Human iPSC-derived hypoimmune cells transplanted into MHC mismatched allogeneic humanized mouse
Having demonstrated the ability of mouse iPSC-derived hypoimmune cells to satisfy each of three testing criteria, the experiments were advanced to evaluate human hypoimmune cells. This was evaluated using a “humanized” mouse system, generated by grafting a functioning human immune system in place of the mouse immune system.
In addition to evaluating the three primary criteria, the ability to successfully engineer human hypoimmune cells from human iPSCs and whether differentiated cells derived from human hypoimmune cells retain biological function were also evaluated.
Creating Hypoimmune Therapeutic Cells from Human iPSCs
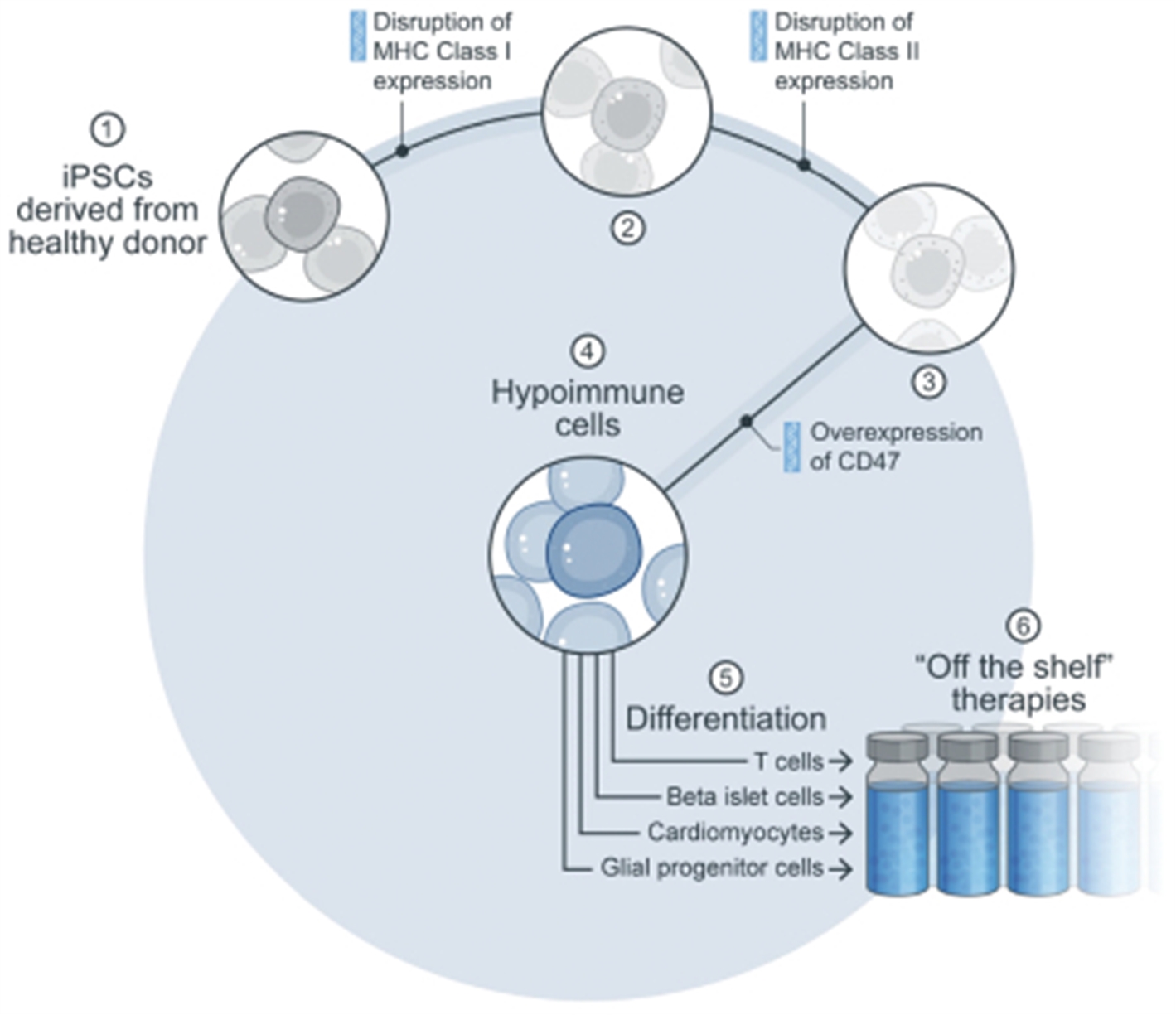
Our hypoimmune technology combines the following three gene modifications to hide cells from the host immune system: Disruption of MHC class I and class II expression (which inactivates adaptive immune responses), and overexpression of CD47 (which hides cells from the innate immune system, including macrophages and natural killer (NK) cells). Pluripotent stem cells from healthy donors are used as the starting material and are then genetically modified with the hypoimmune edits. These edited cells are then differentiated into cell types of therapeutic interest, which are administered to the patient as “off the shelf” therapies.
First, the foregoing three edits were replicated in human iPSCs to engineer a human hypoimmune cell line that had comparable properties to the mouse hypoimmune cells in vitro. Next, non-edited human iPSCs were transplanted into MHC mismatched humanized mice. It was observed that these non-edited human iPSCs were rapidly rejected. Human hypoimmune cells were then transplanted into MHC mismatched humanized mice. It was observed that the human hypoimmune cells survived the full length of the experiment and failed to elicit any type of immune response. From this, it was concluded that, in humanized mice, the human
13
hypoimmune cells can evade the immune system. Pluripotency of human hypoimmune cells was confirmed by differentiation into two different cell types, endothelial cells and cardiomyocytes. These differentiated cells exhibited the characteristics of normal endothelial cells and cardiomyocytes. Finally, to test whether these the differentiated cell types derived from human hypoimmune cells continue to evade the immune system, the differentiated cells were transplanted into humanized mice, and the transplanted cells survived for the full standard observation period. In contrast, differentiated cells derived from non-edited human iPSC cells did not survive after being transplanted, as anticipated. It was also observed that the hypoimmune endothelial cells formed primitive vasculature with active blood flow and the hypoimmune cardiomyocyte cells matured into functional-looking heart cells.
14
Absence of T and B Cell Activation Following Transplantation of Hypoimmune-Edited Human iPSCs into Mismatched Humanized Mice
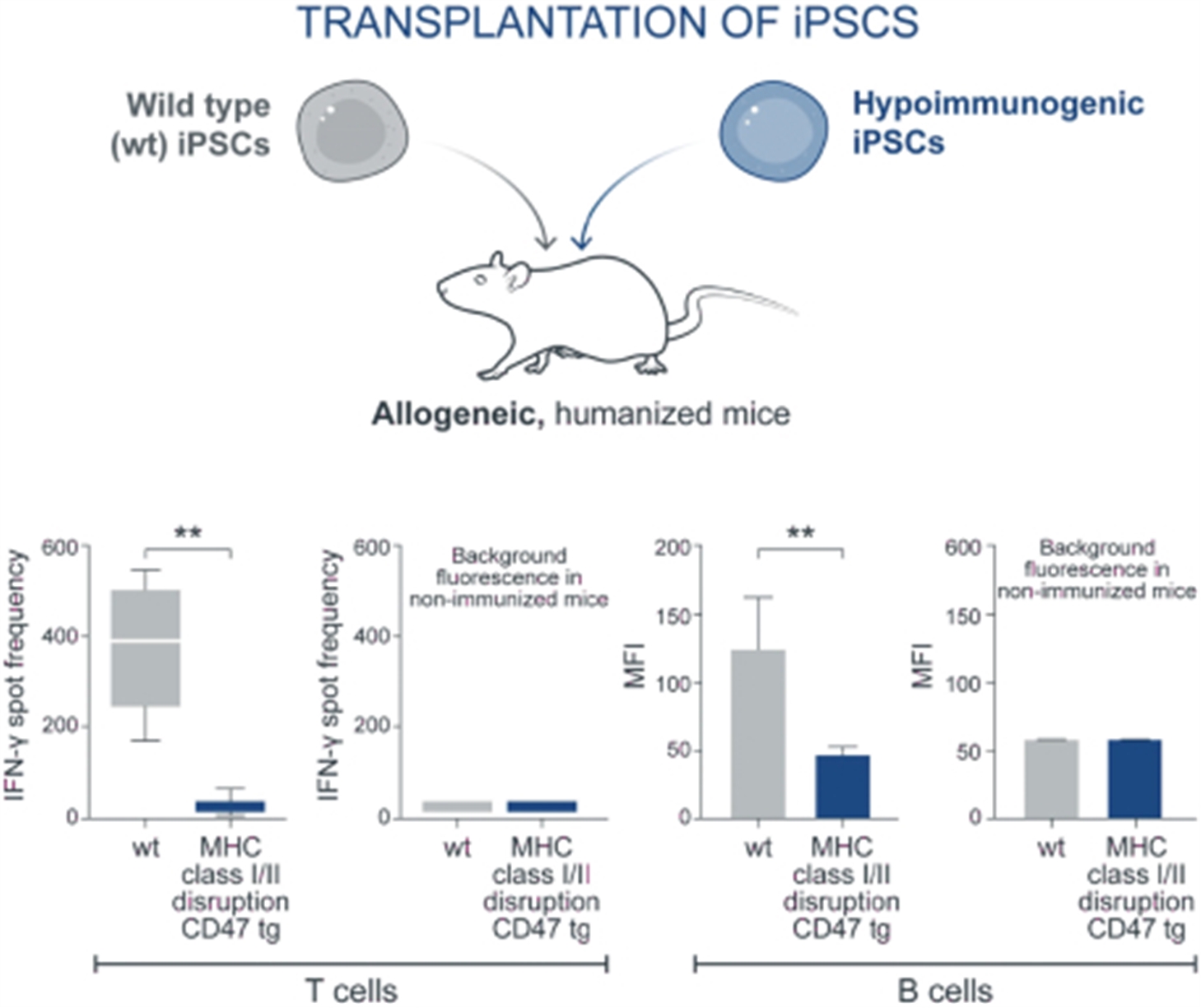
Left panels: T cell activation was measured by EliSpot counts for interferon-gamma production. Immune cells from mice that received wild type (wt) iPSC grafts show a brisk interferon response when tested against allogeneic wt iPSC grafts. In contrast, immune cells from mice that received hypoimmune-edited (MHC class I/II disruption, CD47 tg) cells show only minimal interferon production when exposed to allogeneic hypoimmune cells, comparable to background frequency in non-immunized mice. Right panels: B cell activation was measured by antibody binding to each cell type, shown as mean fluorescence intensity (MFI). Wild type cells exhibit significant antibody binding when incubated with serum from mice that received wt cells. In contrast, hypoimmune-edited cells show only background levels of binding when treated with serum from mice that received hypoimmune-edited cells. Adapted from Deuse et al, Nature Biotechnology 2019.
CD47 is Required to Protect Hypoimmune-Edited Cells from Killing by Human NK Cells
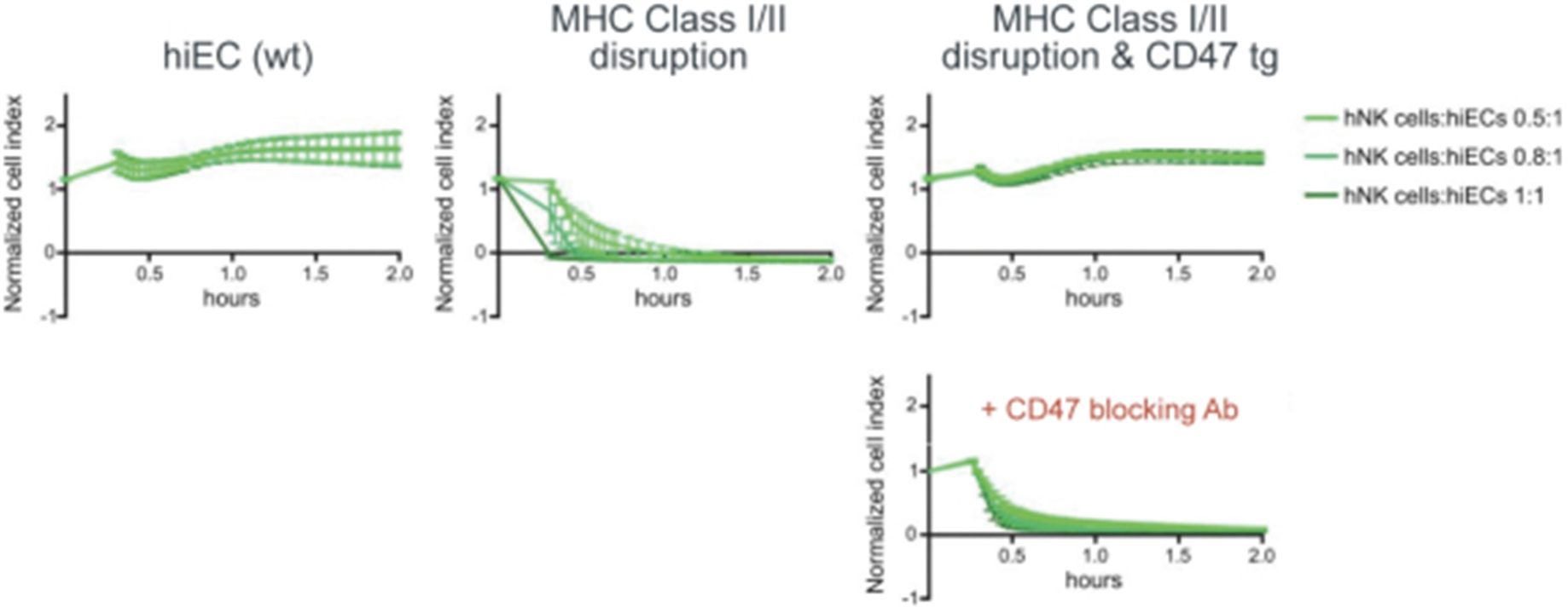
Human iPSCs were differentiated into endothelial cells (hiECs) and plated as a monolayer in a multielectrode system. After exposure to NK cells, monolayer viability was measured electrical impedance, indicated here as normalized cell index. As expected, wt cells were not killed by NK cells. In contrast, cells lacking MHC class I and II (but not expressing CD47 tg; MHC class I/II disruption) were rapidly killed. Addition of CD47 tg prevented killing by NK cells. A blocking antibody to CD47 abolished protection from NK cells, affirming the importance of CD47 overexpression in protection from innate immune cell killing. From Deuse et al, Nature Biotechnology 2019.
15
Survival of Hypoimmune-Edited Human iPSC Grafts in MHC-Mismatched Humanized Mice
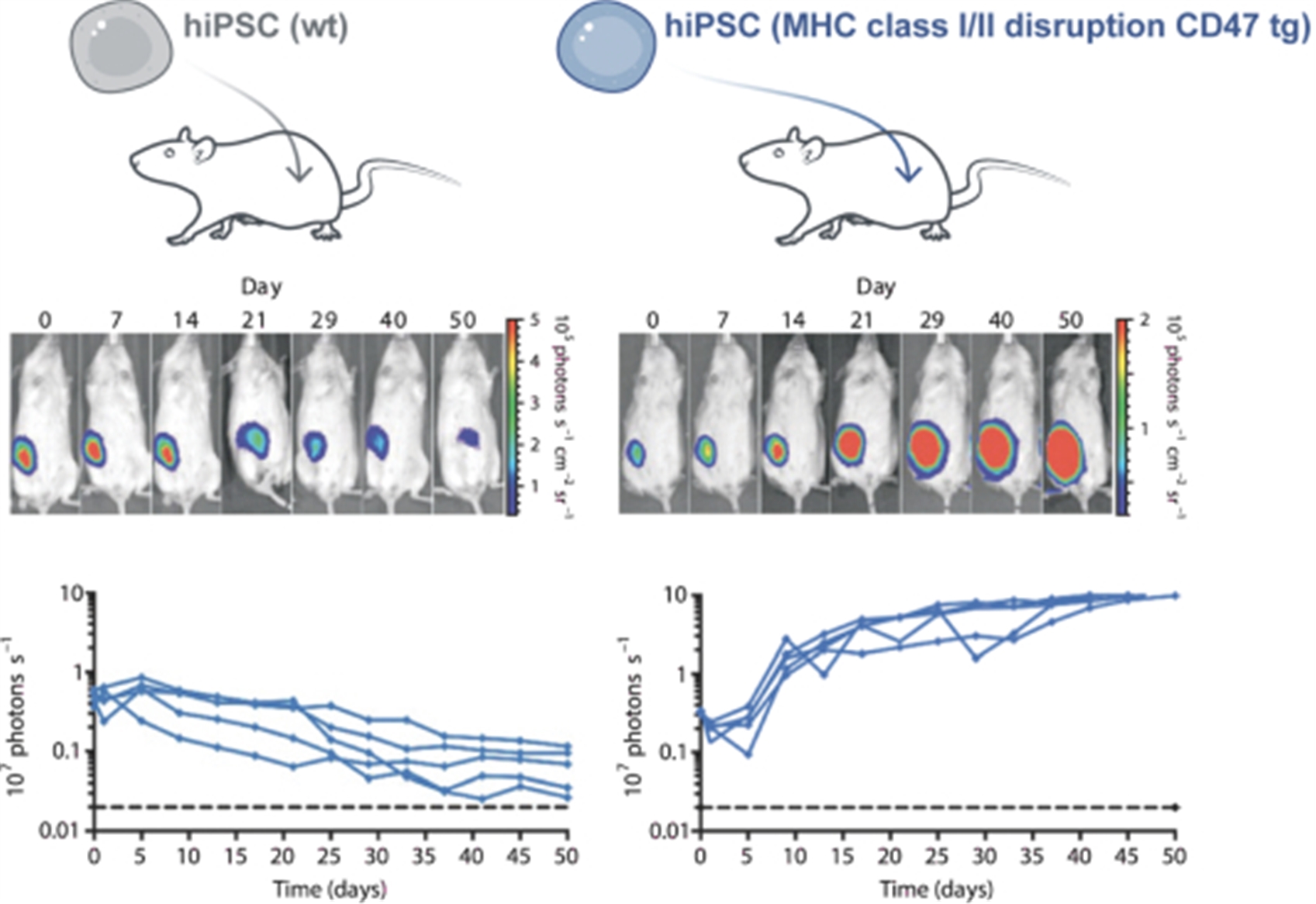
Wild type (wt) and hypoimmune-edited (MHC class I/II disruption CD47 tg) iPSCs were engineered to express firefly luciferase before transplantation. Emission of light was used as an index of graft cell viability. Sequential light emission scans from the same representative animal receiving wt cells show progressive loss of graft viability, indicating graft rejection, confirmed quantitatively in the line tracings below. In contrast, mice receiving hypoimmune-edited cells show graft expansion over the course of the experiment, indicating immune evasion. From Deuse et al, Nature Biotechnology 2019.
NHP hypoimmune cells transplanted into NHPs
To evaluate immune evasion properties of the hypoimmune cells, we have tested the immune response to and survival of hypoimmune iPSCs from NHPs by transplantation into an allogeneic NHP recipient without immunosuppression.
Design for Allogeneic Study Involving Wild Type (Unmodified) and Hypoimmune NHP iPSC Delivery to NHPs
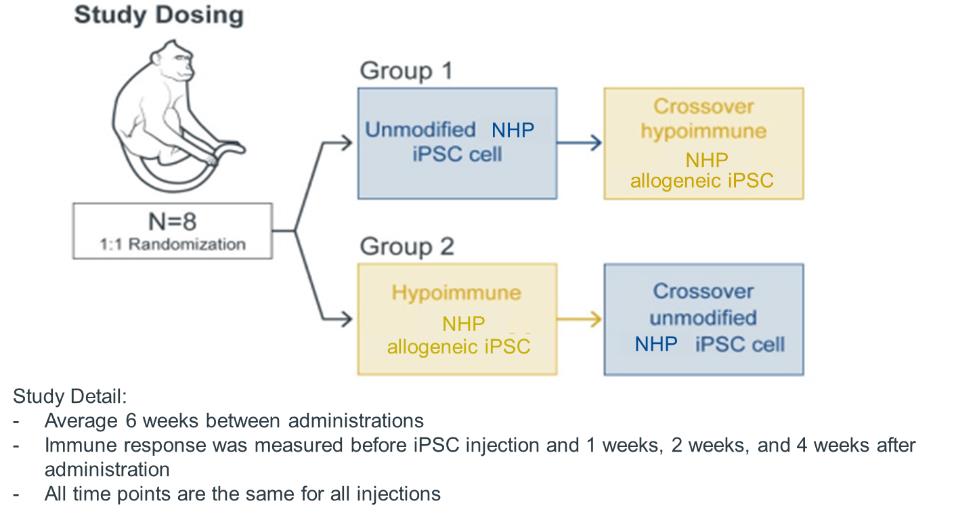
The study involved a randomized group of eight NHPs distributed into two cohorts of four NHPs each. The first cohort received an initial intramuscular injection of non-edited NHP iPSCs and a second injection of NHP hypoimmune cells at six weeks (i.e., a crossover design). The second cohort received an initial injection of NHP hypoimmune cells, which allowed assessment of immune
16
evasion in a naïve recipient. This cohort also received a second injection of non-edited NHP iPSCs, which, with a view towards modeling certain aspects of autoimmune disease, enabled assessment of the impact of injecting hypoimmune cells into an NHP with a pre-existing immune response to non-edited cells. No immunosuppression was administered to any of the animals in the study.
Allogeneic Hypoimmune iPSC Survive in vivo in NHPs with an Intact Immune System
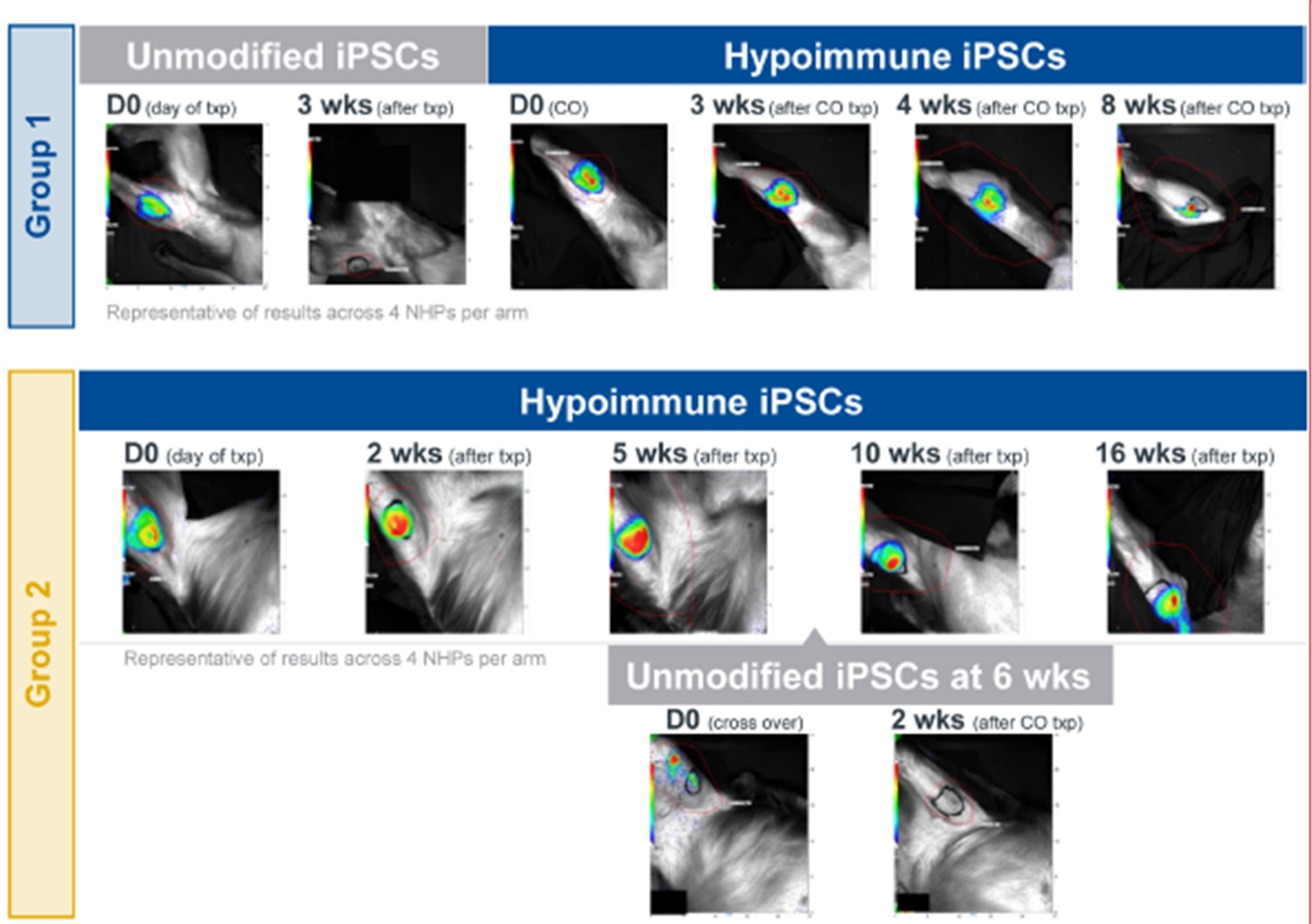
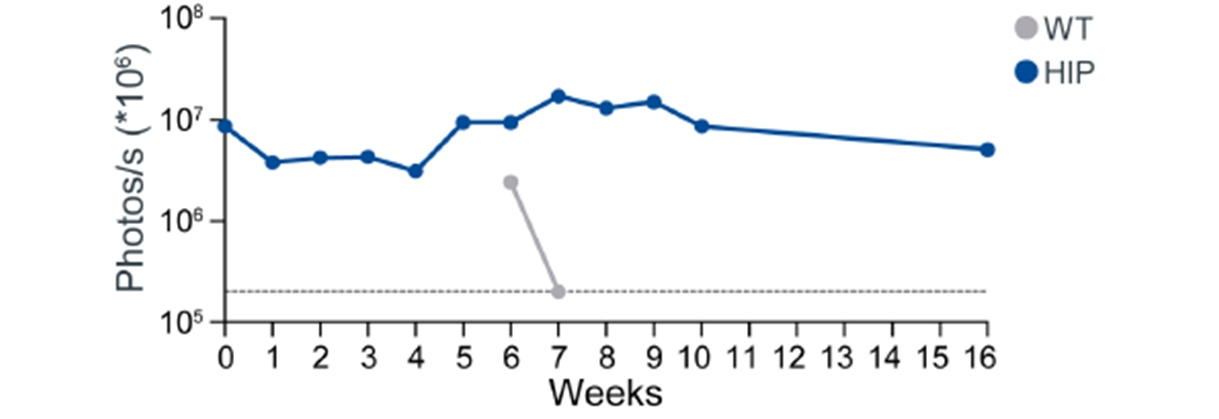
Upper panel: Unmodified wild type (wt) NHP iPSCs (Group 1 top row) or hypoimmune NHP iPSCs (Group 2 bottom row) were introduced via intramuscular injection into allogeneic NHPs. Unmodified NHP iPSCs are undetectable in recipient NHPs by week 3 while hypoimmune NHP iPSCs introduced into naïve NHPs were viable and detectable for 16 weeks post injection. After 6 weeks of the initial injection, NHPs were injected with the cross-over cell type (group 1 with hypoimmune NHP iPSCs and group 2 with wild type iPSCs). In these crossover experiments, hypoimmune NHP iPSCs survived even when the NHP had been exposed to unmodified iPSCs. Unmodified cells injected into NHPs previously injected with hypoimmune iPSCs were rapidly killed with no observable impact on the hypoimmune NHP iPSCs that continued to remain viable. Data are representative for four NHPs receiving HIP iPSCs and wt iPSCs.
Lower panel: iPSC survival is followed over time in vivo using bioluminescence imaging (BLI).
17
Absence of T Cell, B Cell, or NK Cell Responses Following the First Delivery and Crossover of Hypoimmune NHP iPSCs into NHPs
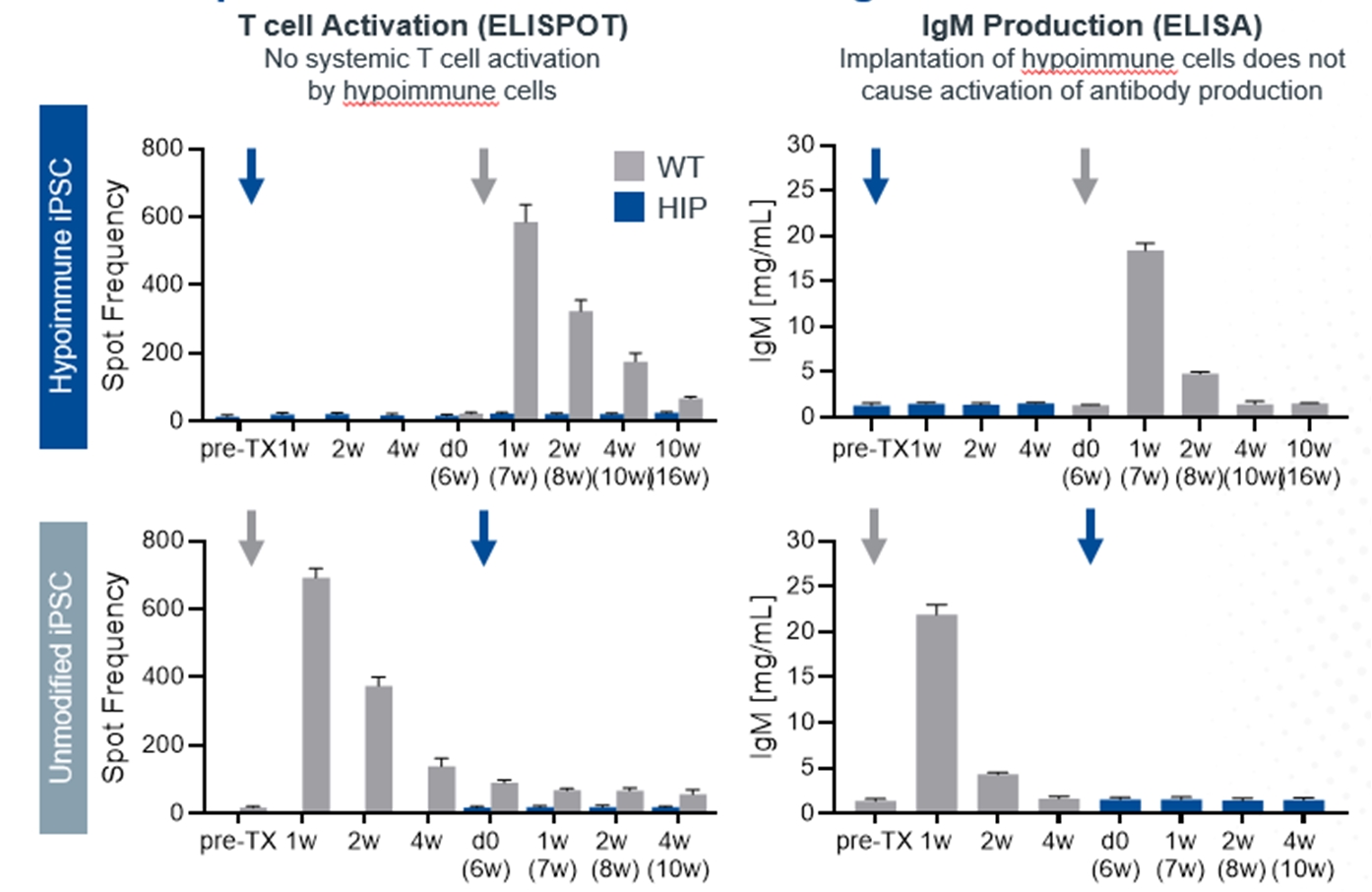

Upper panel: Immune cells from animals receiving hypoimmune iPSCs showed no response when exposed to hypoimmune iPSCs in vitro (Row 1) in contrast to wt iPSCs (Row 2). Lower panel: Neither unmodified nor hypoimmune-edited cells were susceptible to killing by natural killer (NK) cells, indicating protection from the “missing self” signal. Data above are collected from four NHPs in each experimental arm.
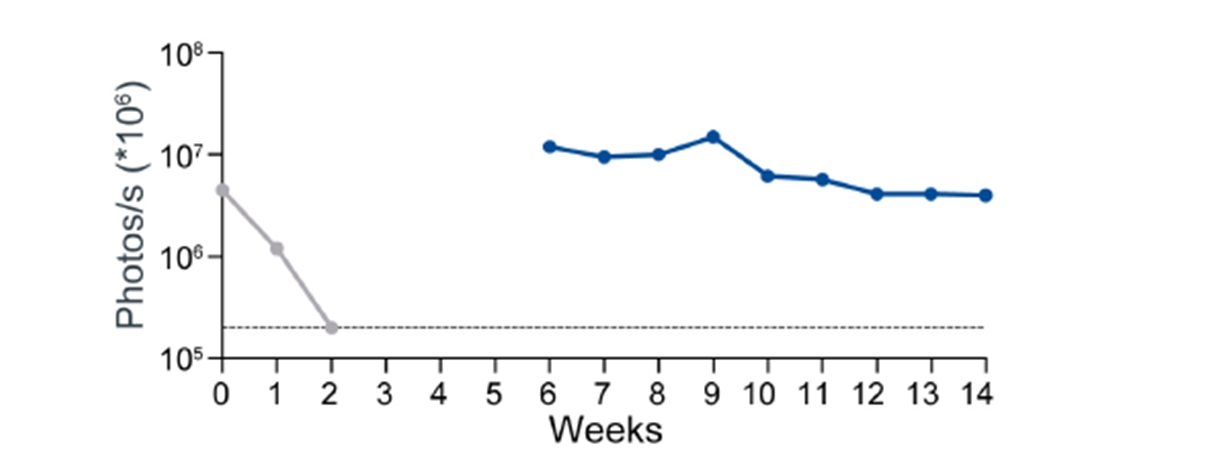
NHP hypoimmune cells grafted into NHPs elicited no detectable systemic immune responses, including no T cell activation and no antibody formation. Innate immune responses mediated by macrophages and NK cells were also undetectable. The transplanted hypoimmune cells were alive and detectable for the duration of the study in these allogeneic recipients (Study duration was 16 weeks for 2/4 NHPs and 8 weeks for 2/4 NHPs. To our knowledge, this is the first instance of prolonged graft survival in an allogeneic transplant setting without immunosuppression in NHPs. In contrast, systemic immune responses from T cells as well as IgM and IgG antibodies were generated to iPSCs without the hypoimmune edits, and the cells were rapidly rejected within two to three weeks.
In the crossover portion of this experiment, injection of NHP hypoimmune cells into NHPs that had previously received non-edited NHPs again elicited no systemic responses as tested in assays for T cell or antibody responses. Similarly, macrophage and NK responses could not be detected. Correspondingly, these cells survived for the full eight weeks that they were monitored suggesting that pre-existing immunity to non-edited human iPSCs had no impact on hypoimmune cell survival. By contrast, in the NHPs that had previously been injected with the hypoimmune cells, the non-edited NHP cells elicited both T cell and antibody responses against the non-edited cells. Notably, these non-edited cells were rapidly rejected (in one to two weeks) in the recipient even as the previously injected hypoimmune cells continued to be viable in the other leg of the NHP. These results provide confirmation that the survival of the hypoimmune allo-graft was not an artifact of an impaired immune system or immune response in the recipient NHP. They also suggest that these hypoimmune cells have the potential for immune evasion even the context of a new immune response toward cells without these edits.
18
In light of our preclinical data to date, we believe our hypoimmune technology has the potential to address the most fundamental limitation of ex vivo therapies, persistence, and thereby unlock waves of potentially disruptive therapies across a variety of cell types.
Safety Switch for Hypoimmune Cells
We are actively investigating approaches to control hypoimmune cells after administration into the patient. If necessary, the aim of these “safety switches” would be to provide a mechanism to eliminate hypoimmune cells within the body in a targeted fashion, in scenarios where the cells are not in a location where physical removal is viable. Such a safety switch would be beneficial to mitigate the potential risk of a hypoimmune cell becoming infected with a virus or undergoing oncogenic transformation, in light of the immune evasion modifications to these cells.
One approach we are exploring as a safety switch is to re-sensitize the hypoimmune cells to innate cell killing via administration of a blocking anti-CD47 antibody. We have tested the effectiveness of this approach in iPSCs and teratomas (a particular tumor formed by pluripotent cells with histological features from all three germ layers), both bearing the hypoimmune modifications. Using hypoimmune NHP iPSCs we observed in vitro that the addition of an anti-CD47 antibody binds to and blocks CD47 expressed in the hypoimmune cells and restores the sensitivity to the missing-self killing response mediated by NK cells. We also assessed this strategy in mouse experiments, where the animals were transplanted in with human iPSCs which then formed small teratomas. We have shown that treatment with an anti-CD47 antibody resulted in the loss of immune evasion and the rapid killing of these transplanted cells. We have identified several additional safety switches with in vivo activity and intend to continue to explore them, potentially including multiple safety switches in therapeutic programs moving forward.
Anti-CD47 Administration Results in the Rapid Clearance of Hypoimmune NHP iPSCs in vitro
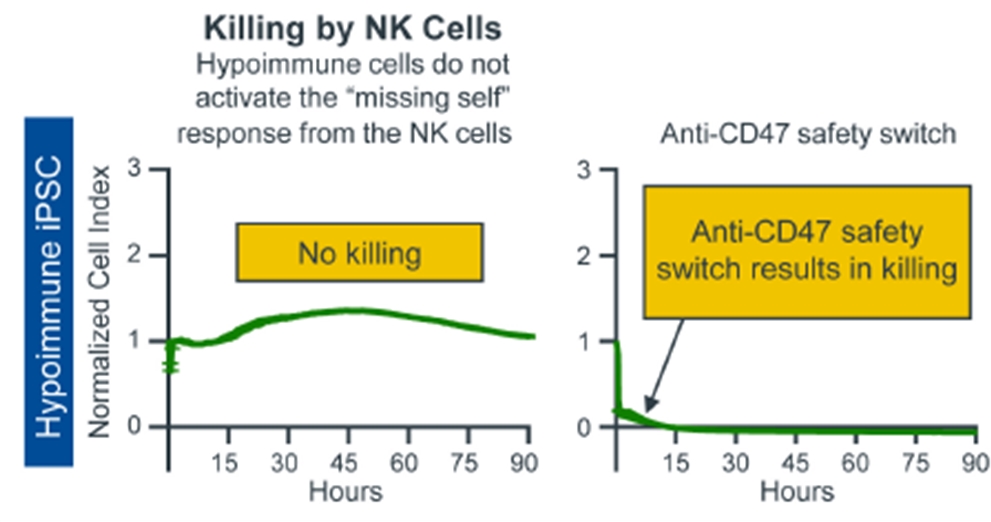
On left: Hypoimmune NHP iPSCs do not induce killing by NK cells in an in vitro killing assay; On right: by contrast, Anti-CD47 antibody treated hypoimmune NHP iPSCs are no longer able to evade missing-self responses mediated by NK cells and are killed rapidly.
Anti-CD47 Administration Results in the Rapid Clearance of Human iPSCs derived Teratomas in a Humanized Mouse Model
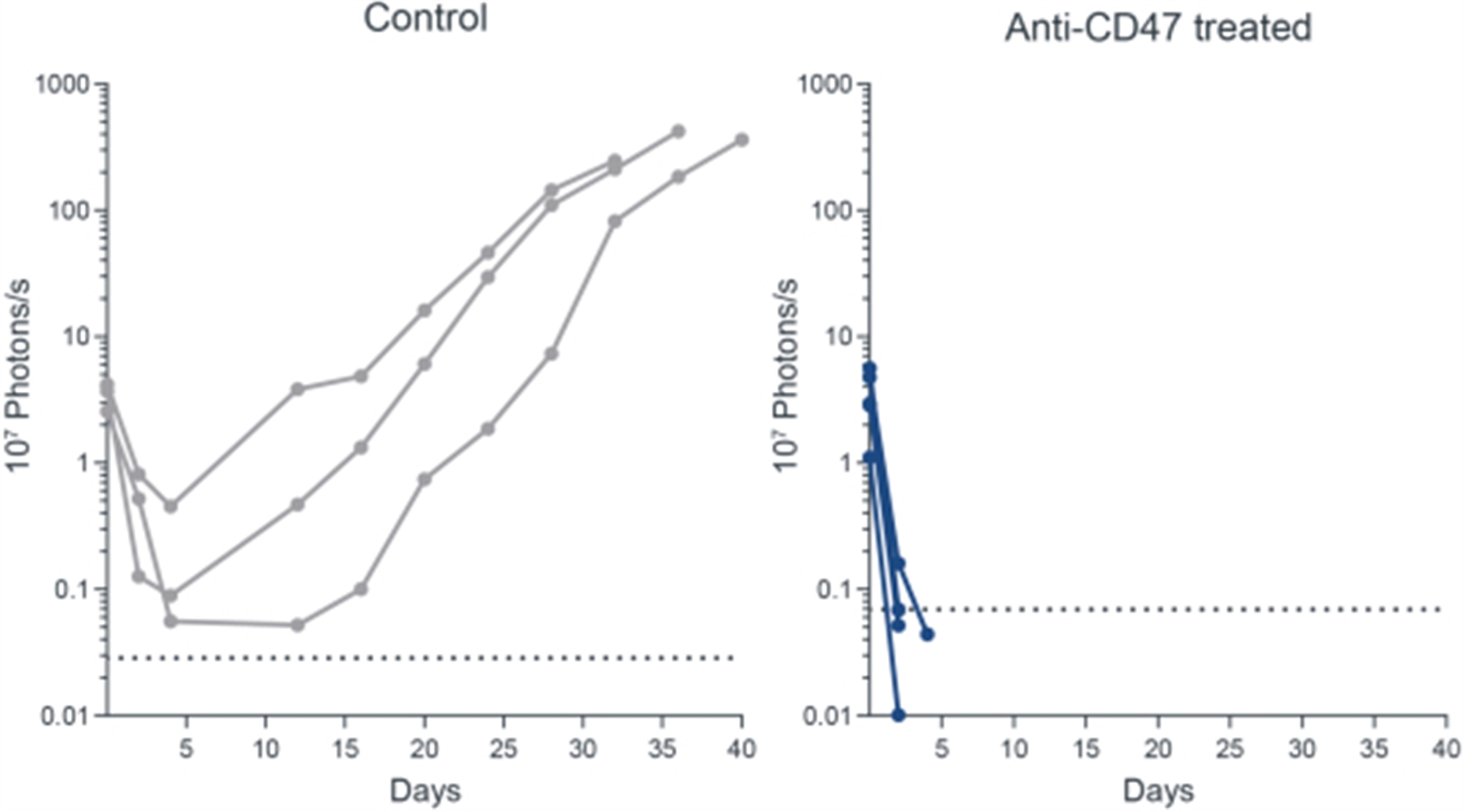
19
On left: Human iPSC proliferate (as visualized by luminescence of live cells) and form teratoma in NSG mice (n=3) with adoptive transferred human NK cells. Administration of isotype control has no impact on HIP survival. On right: Blocking of CD47 in vivo results in killing of HIP iPSCs (as visualized by luminescence of live cells) in NSG mice (n=5) with adoptive transferred human NK cells.
CD47 overexpression is differentiated in inhibiting “missing self” response relative to other approaches
As part of our ongoing program to further refine our hypoimmune technology we evaluated the effectiveness of the overexpression of CD47 in comparison to other molecules that have at least some ability to inhibit innate immune responses. We carried out these head-to-head comparisons in K562 cells, a naturally MHC class I and class II deficient cell line. The lack of the MHC class I molecule should result in virtually instantaneous cell killing by stimulated innate immune cells such as NK cells due to the activation of the “missing self” response. We compared three molecules, HLA-E, HLA-G and PDL-1, that have previously been proposed to have a role in inhibiting innate immune responses versus CD47. In this assay, overexpression of these three molecules conferred limited protection from NK cell killing in contrast to CD47 overexpression. This difference in activity may be the result of the more ubiquitous presence of the receptor for CD47 receptor on innate immune cells relative to the presence of receptors for these other immunomodulators. While these results do not rule out a role for these other molecules in inhibiting NK cell responses, they suggest that CD47 may be sufficient to nullify the NK cell-mediated missing-self response.
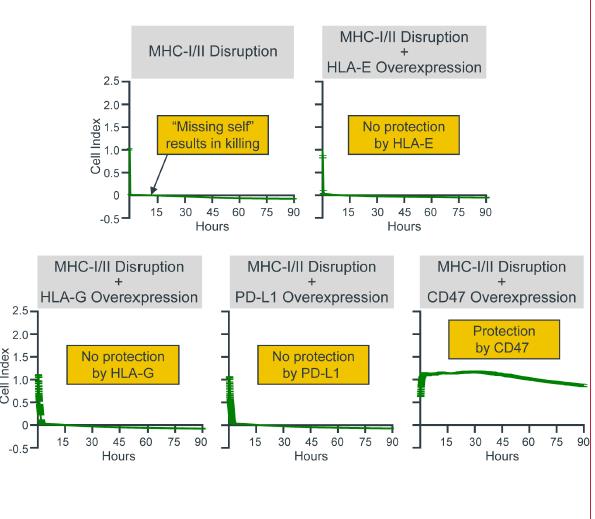
Panels above show in vitro killing assays mediated by NK cells. Cells missing MHC molecules are killed by NK cells, as measured by rapid decline in cell index. Overexpression of immunomodulatory molecules such as HLA-E, HLA-G or PDL-1 in cells missing MHC molecules did not block NK cell killing. By contrast, overexpression of CD47 blocked NK cell mediated “missing-self” response.
Our ex vivo Cell Engineering Pipeline
Allogeneic T Cell Program (SC291, SC276, SC255)
Our allogeneic T cell program utilizes T cells from healthy donors to generate CAR T therapies that will initially target CD19, a protein expressed on the cell surface of B cell malignancies, to treat patients with refractory lymphoma. We believe that applying the hypoimmune technology to allogeneic T cells gives us an opportunity to create differentiated allogeneic CAR T therapies.
We believe our allogeneic T cell and T cell fusosome discovery programs provide us with two potentially disruptive programs to address the limitations of adoptive T cell therapy for cancer, each with idiosyncratic risks and opportunities. We also believe each approach can address separate and valuable opportunities if they are both successful. Specifically, our allogeneic T cell program offers the opportunity to perform multiple gene edits in a T cell, which may allow us to make intentional modifications to control T cell function or to deliver more complex chimeric receptors and signal integration machinery to enable the T cell to distinguish tumor cells based on surface antigen combinations to improve the specificity of targeting. These approaches may prove especially valuable in
20
targeting solid tumors, which have remained largely refractory to CAR T approaches to date. We also have an earlier-stage program looking to differentiate hypoimmune iPSCs into T cells. While we are still working to successfully create the appropriate T cells from an iPSC, we expect that progress with our allogeneic T cell program will also inform the iPSC T cell program. Separately, the fusogen technology allows for the in vivo generation of CAR T cells in a patient, offering a distinct advantage in terms of manufacturability and scalability that may enable the introduction of gene-modified T cells earlier in the course of a patient’s therapy. Additionally, modifying the T cells inside the body without the need for ex vivo manipulation of the cells may generate CAR T cells with more favorable attributes.
We intend to develop our CD19 allogeneic T cell therapies with the goal of submitting an IND for SC291 in 2022. We intend to follow this with SC276, a CD22-targeting allogeneic CAR T which could be combined with targeting CD19 and offers the potential benefit of higher and more durable complete response rates, with an IND submission as early as 2023. We are also advancing our SC255 allogeneic T cell program targeting BCMA for multiple myeloma, with the goal of submitting an IND in the next several years.
Background on B cell Malignancies
B cell malignancies represent a spectrum of cancers including non-Hodgkin Lymphoma (NHL), chronic lymphocytic leukemia (CLL), acute lymphoblastic leukemia (ALL), and multiple myeloma (MM) and result in over 100,000 deaths per year in the United States and Europe. See the subsection titled “—in vivo Cell Engineering Pipeline—Background on B Cell Malignancies” for further background discussion. In addition to our in vivo cell engineering technology, we believe our ex vivo cell engineering technology also has the potential to address B cell malignancies.
Current Treatment Landscape and Unmet Need
We believe our hypoimmune edited cells have the potential to create a differentiated platform for developing allogeneic T cells. There are two major hurdles to the use of allogeneic T cells. The first is the risk of graft versus host disease, in which the allogenic donor T cells target and kill recipient tissues. Multiple CAR T cell product candidates in clinical development have managed to prevent this reaction through gene edits targeting components of the T cell receptor such as TCR-alpha gene. The more significant challenge has been HvGR, in which the patient’s immune system kills the transplanted T cells. One strategy to approach this challenge has been to essentially eliminate the patient’s immune system, neutering its ability to find and destroy the transplanted allogeneic CAR T cells. This strategy has two limitations. First, the patient is at risk of severe infections during this period of substantial immune suppression. Second, as the immune system returns, it will inevitably reject the allogeneic CAR T cells, limiting the duration that these therapeutic cells are in the body. Experience with autologous CAR T cells in patients with B cell malignancies has demonstrated that persistence of CAR T cells is important for the durability of response. Thus, the ability to effectively prevent long term rejection of an allogeneic CAR T therapy without significant immune suppression would be a major advance. We are aware of other efforts to develop allogeneic CAR T cell products that focus on overcoming the adaptive immune system (T and B cells). Our technology addresses rejection mediated by both the adaptive and innate immune systems, giving us the potential to create a differentiated allogeneic CAR T solution.
Our Allogeneic T Cell Program Approach
Our hypoimmune technology is designed to hide the cell from the patient’s immune system, and we are applying this technology to manufacture allogeneic CAR T cells. We intend to utilize T cells from healthy donors into which we will introduce the CAR gene and make the gene modifications necessary to overcome graft versus host disease and to incorporate our hypoimmune technology in an effort to address host versus graft response. We then intend to expand these cells ex vivo, with a goal of making many batches from a single donor as well as creating comparable CAR T cells from various healthy donors. These allogeneic CAR T therapies could be frozen and delivered as an “off the shelf” product for cancer patients without the need for severe immunosuppression.
Preclinical Data
For our preclinical studies, human donor-derived T cells were genetically modified ex vivo, to generate cells bearing the hypoimmune edits (disruption of MHC class I/class II; overexpression of CD47), TCR-alpha disruption (to mitigate graft versus host disease) and the expression of a CD19 CAR. These cells were then tested in vivo for their tumor-killing activity in a human xenograft mouse model for leukemia (Nalm-6). These preclinical data suggest that the hypoimmune edits do not interfere with CAR T killing activity. We observed clearance of the leukemic cells by the hypoimmune CD19 CAR T cells and the potency of these cells was comparable to unmodified CD19 CAR T cells, which are similar to CAR T cells currently in clinical use.
21
Hypoimmune Donor-Derived CD19 CAR T Cells Clear Leukemia Cells in a Human Xenograft Mouse Model at Levels Comparable to Unmodified CD19 CAR T Cells
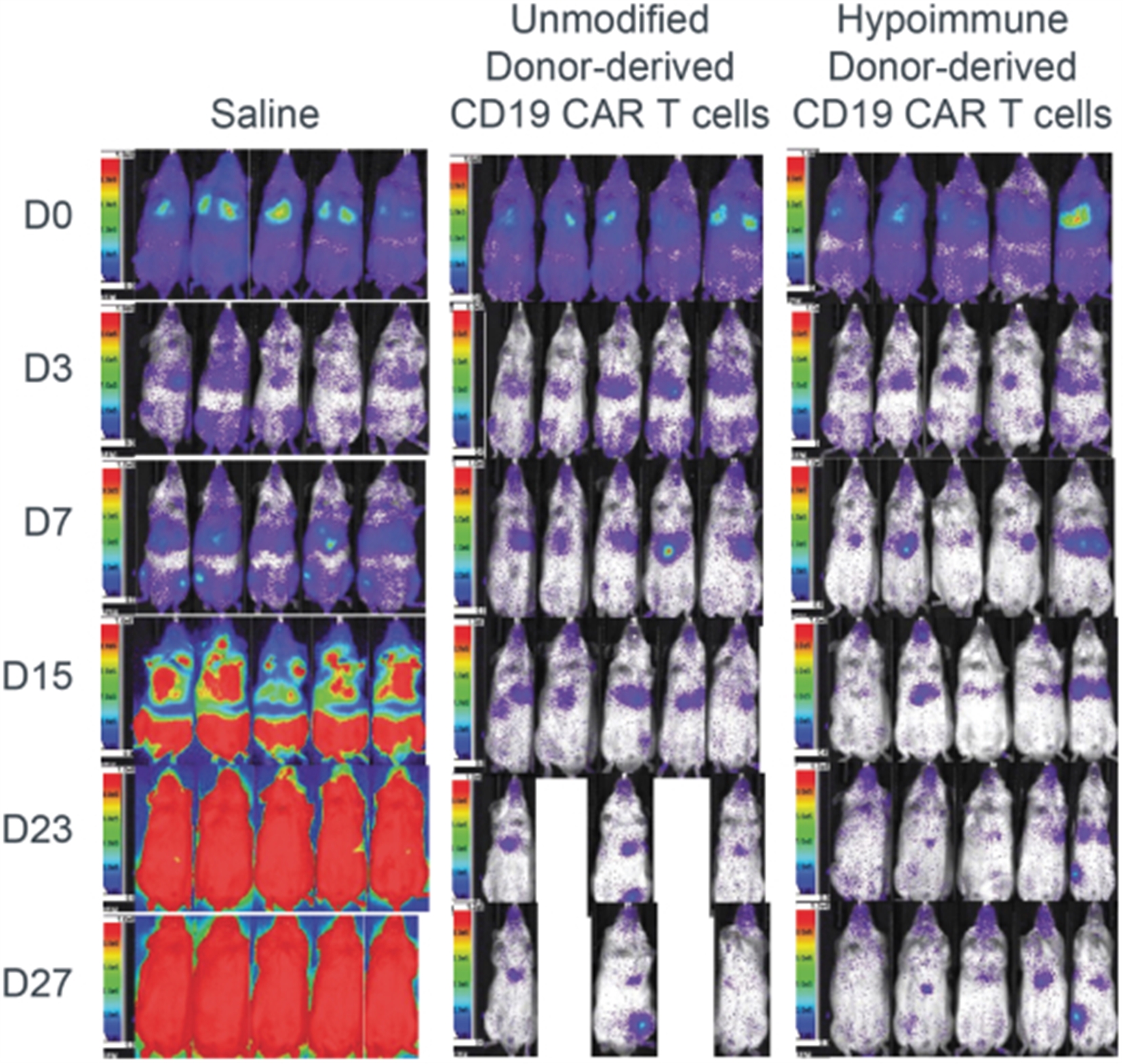
Activity of hypoimmune donor-derived CD19 CAR T in a mouse leukemia xenograft model (Nalm-6). Note that when compared to untreated controls, infusion of unmodified CD19 CAR T or hypoimmune CAR T results in eradication of leukemia cells. Both cohorts of CAR T treated mice had significantly reduced tumor burden when compared to control as early as D7 (p≤0.0001; One-way ANOVA Bonferroni) with no significant difference between either of the treatment arms.
22
Furthermore, the hypoimmune CD19 CAR T cells were protected from immune system rejection in humanized mice, and no evidence was observed of either adaptive or innate immune system activation, in contrast to the unmodified CAR T cells:
Absence of T Cell, B Cell, NK Cell, and Macrophage Responses in a Human Xenograft Mouse Model Following Injection of Hypoimmune Donor-Derived CD19 CAR T Cells
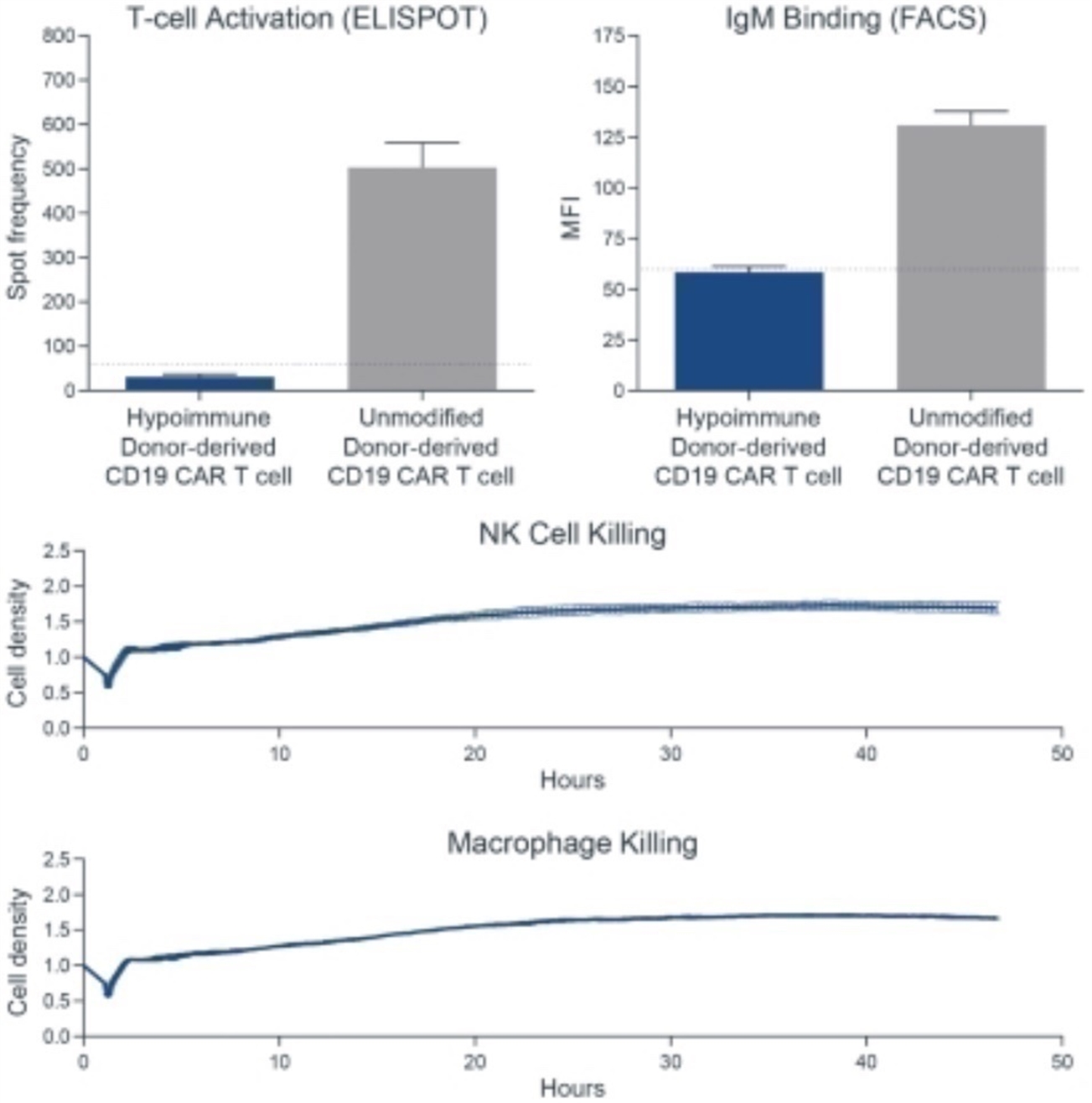
Immune cells from humanized animals receiving hypoimmune donor-derived CD19 CAR T showed no response when exposed to hypoimmune CAR T cells in vitro. Delivery of hypoimmune cells did not result in production of donor-specific antibodies, as evidenced by binding of IgM antibodies to the surface of donor cells (increased mean fluorescent intensity, MFI). Hypoimmune-edited cells were not susceptible to killing by NK cells nor macrophages, indicating protection from the “missing self” signal.
Development Plan and Key Next Steps
Process development work is ongoing within the Technical Operations team to develop scalable manufacturing processes to generate quality and consistent allogeneic T cell product candidates. In parallel, our cell engineering team is developing an efficient and specific gene editing platform to enable manufacturing of our allogeneic T cell product candidates.
The next major milestones are to complete Good Laboratory Practice (GLP) studies and GMP manufacturing with the goal of submitting an IND for SC291 in 2022. We believe that early data from the SC291 study will help us understand the therapeutic potential of this therapy, as the ability to evade immune detection with corresponding enhanced persistence of the CAR T may predict higher and more durable complete responses for patients. Additionally, it will give us insight into the potential of this hypoimmune platform for additional CAR T programs targeting other antigens and cancers as well as the potential for the platform more broadly in areas beyond cancer. Following the IND submission for SC291, we are progressing SC276, a CD22-targeting allogeneic CAR T which could be combined with targeting CD19 and offers the potential benefit of higher and more durable complete response rates, with the goal of submitting an IND as early as 2023. We are also advancing our SC255 allogeneic T cell program targeting BCMA for multiple myeloma, with the goal of submitting an IND in the next several years.
Beta Cell Program
Our beta cell program aims to restore lifelong glucose control in Type I diabetes mellitus (T1DM), patients by transplanting hypoimmune iPSC-derived beta cells. Current therapies for T1DM require continual management, and we believe that effectively restoring beta cell functionality will meaningfully improve patient outcomes for patients with T1DM. We intend to develop this program with the goal of submitting an IND for SC451 as early as 2023.
23
Background on Type 1 Diabetes
T1DM is an autoimmune disease in which the patient’s immune system destroys its own pancreatic beta cells. The destruction of these cells leads to complete loss of insulin production and a metabolic disease wherein patients are unable to control their blood glucose levels. Often called “juvenile diabetes”, this disease commonly has its onset in adolescence. Beta cells reside in specialized hormone-producing clusters within the pancreas called the Islets of Langerhans. In T1DM, activated T lymphocytes infiltrate the islets and selectively kill the beta cells, progressively reducing the body’s capacity to produce insulin. Once the reserve capacity of beta cells is exhausted, blood glucose rises, and the patient will have a life-long battle to control blood glucose levels.
T1DM affects 1.6 million adults in the United States, and there are approximately 20,000 new cases diagnosed per year in patients under the age of 20. In Europe there are an estimated 2.4 million adults with T1DM, and 300,000 under the age 20, with 31,000 new cases of T1DM diagnosed each year. Combining prevalence in the United States and Europe yields a pool of approximately 4 million patients with T1DM.
Current Treatment Landscape and Unmet Need
Insulin injection is the main treatment option for T1DM. Despite significant advances in types of insulins, glucose monitoring, and insulin pumps, life expectancy for T1DM is still approximately 15 years shorter than for people without diabetes. Patients are at risk from acute complications of hyperglycemia, including diabetic ketoacidosis and coma. Conversely, they are also at risk of hypoglycemic episodes, particularly at night, which can lead to the “dead in bed” syndrome, thought to result from cardiac arrhythmias induced by low glucose. Long term elevations in blood glucose levels have particularly devastating effects on arteries and capillaries, resulting in premature myocardial infarction, stroke, limb ischemia, gangrene, kidney failure, and blindness due to diabetic retinopathy. “Insulin pumps,” which feature a computerized system for sensing blood glucose and delivering appropriate doses of insulin, have improved glycemic control. Notably, data from the FDA indicate that issues with insulin pumps are among the most frequently reported problems in their database. All current therapies require patients to carefully monitor their dietary intake, which, while inconvenient in adults, is a frequent point of failure in adolescents.
Pancreas transplantation for uncontrollable diabetes was first performed in the 1960s, and this established the principle that replacing the beta cells (here in the context of the whole pancreas) could restore physiological glucose control. Pancreas transplants are complicated surgical interventions, require lifelong immunosuppression, and are limited due to organ availability. Nevertheless, some 30,000 pancreas transplants have been performed worldwide to date.
Because of these challenges, the biomedical community began exploring pancreatic islet transplantation in the 1970s. This process involves enzymatic digestion of a donor pancreas and isolation of the Islets of Langerhans followed by delivery of these cells to an appropriate site in the body where the islets can engraft and become well vascularized. The major lessons from islet transplantation have been that glucose homeostasis can be restored, insulin-independence can be achieved, hemoglobin A1C levels (a marker of long-term glucose levels) can be normalized, and severe episodes of hypoglycemia can be reduced. As with an organ transplant, patients must be immune suppressed to prevent immune rejection of the transplanted cells. In addition to complications from this immune suppression and the lack of cell availability, the principal limitation of islet transplantation has been the therapy’s durability. Most patients lose glucose control over months to years and eventually become insulin-dependent again, primarily due to immune rejection of the allogeneic islets.
Our Beta Cell Program Approach
The goal of our beta cell hypoimmune program is to restore lifelong glucose control in T1DM patients by transplanting hypoimmune iPSC-derived beta cells, including beta cells. Our goal is to create a therapy that restores the body’s normal beta cell mass, giving patients physiologically appropriate glucose sensing ability and insulin secretion. We believe this therapy could reduce, or even eliminate, the hypoglycemia and hyperglycemia in diabetic patients, potentially enabling less onerous and costly treatment, fewer complications, and longer life expectancy, resulting in a meaningfully improved quality of life.
We focus our efforts around three goals: (i) deriving highly functional beta cells from PSCs, (ii) genetically modifying these cells to evade allogeneic immune responses, and (iii) genetically modifying these cells to evade autoimmune destruction of beta cells. This strategy requires building on lessons from pancreatic islet transplantation, recent advances in understanding pancreatic islet developmental biology, and our hypoimmune technology.
Deriving beta cells from iPSCs has the potential to solve limitations associated with donor pancreas and improve the overall product quality and product consistency. iPSCs have the potential to create a virtually limitless supply of these cells. Our program uses proprietary differentiation protocols to generate mature beta cells with glucose control comparable to primary human islets, as evidenced by our animal studies. Finally, we intend to modify the genomes of the iPSCs in order to apply our hypoimmune technology. If successful, the hypoimmune gene modifications will protect these cells from both auto-immune and allogeneic rejection
24
by the patient’s immune system and potentially remove the need for toxic immunosuppression in transplant recipients. Hypoimmunity also should eliminate the need for physical separation of the beta cells from the rest of the body by a device or encapsulation technology, which may allow for tighter control of glucose by eliminating the lag time between glucose sensing and insulin secretion.
Preclinical Data
We are developing a proprietary protocol that will differentiate hypoimmune iPSCs into mature, glucose-sensitive, insulin-secreting beta cells based on licensed technology from Washington University in St. Louis. This technology enables differentiation of beta cells at a greater purity and with superior function compared to published stem cell-based protocols. The principal function of beta cells is to maintain steady levels of glucose in circulation. The beta cells sense when glucose levels rise in the bloodstream and release insulin in response. In vitro, our beta cells respond to glucose and robustly secrete insulin at an equivalent level to primary human islets, as depicted in the figure below.
Human iPSC-Derived Beta Cells Exhibit Glucose-Induced Insulin Release
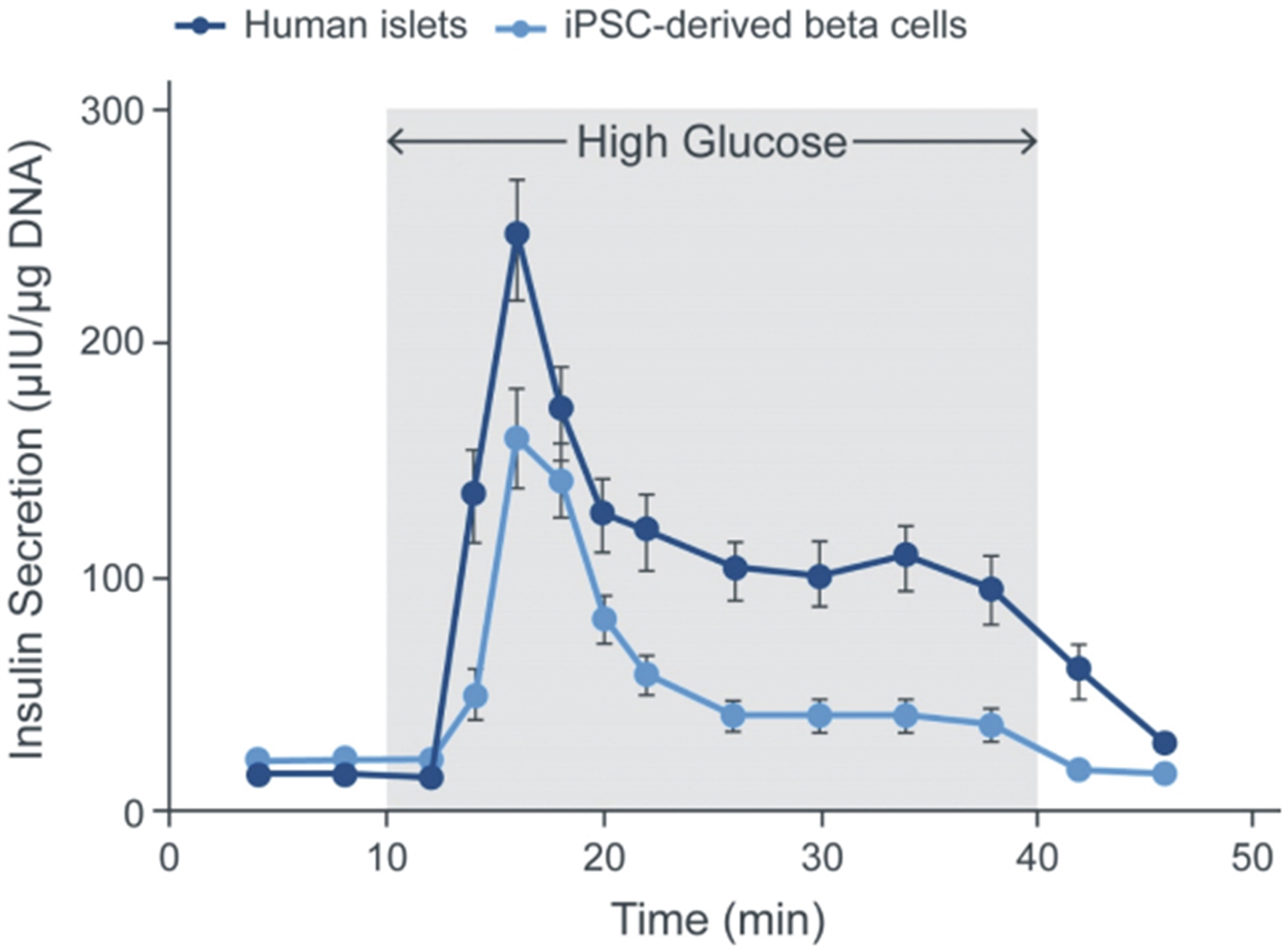
Human islets from cadaveric pancreases (gold standard) exhibit robust insulin secretion in response to an increase in glucose levels. Human iPSC-derived beta cells using technology licensed from Washington University in St. Louis show similar dynamics of insulin secretion to the cadaveric islets.
These stem cell-derived beta cells were tested in a mouse model of Type I diabetes induced by the beta cell toxin, streptozotocin. When transplanted into the kidney of the diabetic mice, these beta cells normalize glucose levels in an equivalent fashion to primary human islets. The diabetic glucose levels return when the grafts are surgically excised (nephrectomy). Similar to the human phenotype, diabetic mice cannot normalize circulating glucose levels following a glucose injection. Following transplantation of our beta cells, these mice rapidly normalized blood glucose in an equivalent fashion to both non-diabetic mice and diabetic mice that received human primary islet transplants.
25
In vivo Performance of iPSC-Derived Beta Cells in a Mouse Model of T1DM
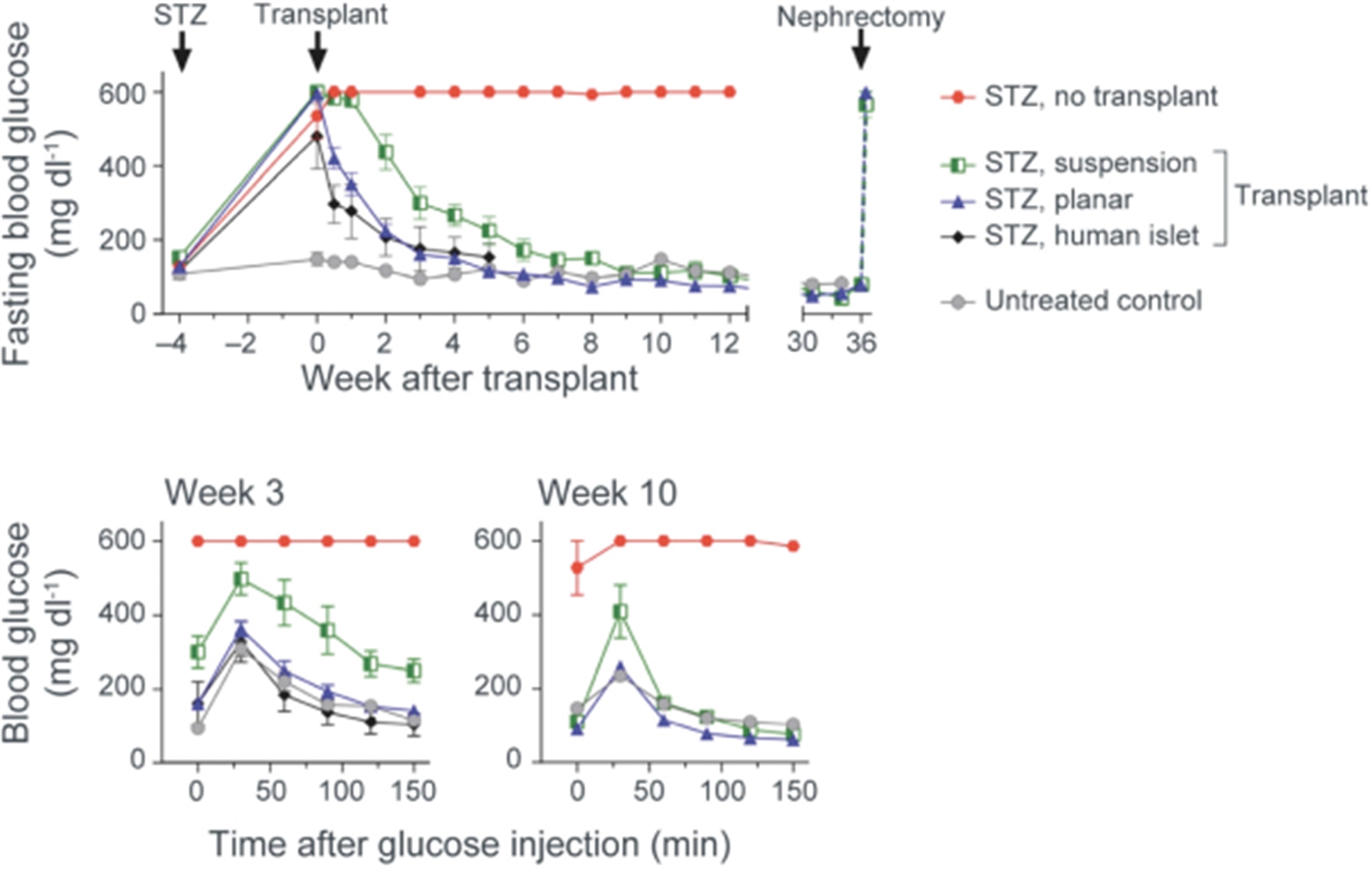
Top: Normalization of blood glucose levels after transplantation of cadaveric human islet cells or iPSC-derived islet cells obtained by planar or suspension differentiation (Washington University technology). Note the rapid normalization of blood glucose with cadaveric and stem cell-derived islets with the planar protocol, with slower normalization using the suspension protocol. In all groups, removal of the graft by nephrectomy re-induced diabetes, indicating the correction resulted from the transplant. STZ, streptozotocin, is a toxin for beta-islet cells that induces diabetes in animal models. Bottom: Normalization of blood glucose after glucose injection by transplantation of cadaveric islet cells or iPSC-derived islet cells. Note the more complete normalization using the planar protocol. Groups are defined by the same symbols shown in the middle panel. From Hogrebe et al, Nature Biotechnology 2020.
We ran an experiment to better understand whether hypoimmune edits impair the function of islet cells and to confirm that these edits enable the islets to evade immune responses. For these experiments, we made the hypoimmune genetic modifications in human islets isolated from cadaveric human donors. These cells were transplanted intramuscularly, without immunosuppression, into a humanized mouse strain, a mouse strain engrafted with human immune cells that mimic certain aspects of the human immune system. Type 1 diabetes was induced in this mouse model with streptozotocin, and the ability of transplanting islet cells to restore normal glucose levels while evading immune rejection was tested. We found that hypoimmune edited human donor islets were successful in restoring glucose control while unmodified islets failed to do so. The key driver of this difference was that, while human donor islets with hypoimmune edits survived for the full duration of the experiment (approximately 30 days), unmodified islets were rapidly rejected within a week. These data demonstrate that hypoimmune edits do not impair the ability of cadaveric islet cells to restore glucose homeostasis, but they do prevent rejection of these cells in mice with humanized immune systems.
We next tested whether our hypoimmune edited cells could survive in serum containing autoreactive antibodies and T cells from patients with Type I diabetes. We exposed pancreatic islets with or without our hypoimmune edits to serum and T cells from patients with Type I diabetes. As expected, both T cells and antibodies from patients recognize and kill unedited pancreatic islets. In contrast, these T cells and antibodies from Type I diabetics did not recognize or kill our hypoimmune edited islet cells. The results from this experiment increase our confidence that, in addition to preventing allogeneic rejection, our hypoimmune technology may allow islet cells to survive autoimmune killing in patients with Type I diabetes without the need for immunosuppression.
26
Hypoimmune human donor pancreatic islet cells evade immune detection and regulate glucose levels
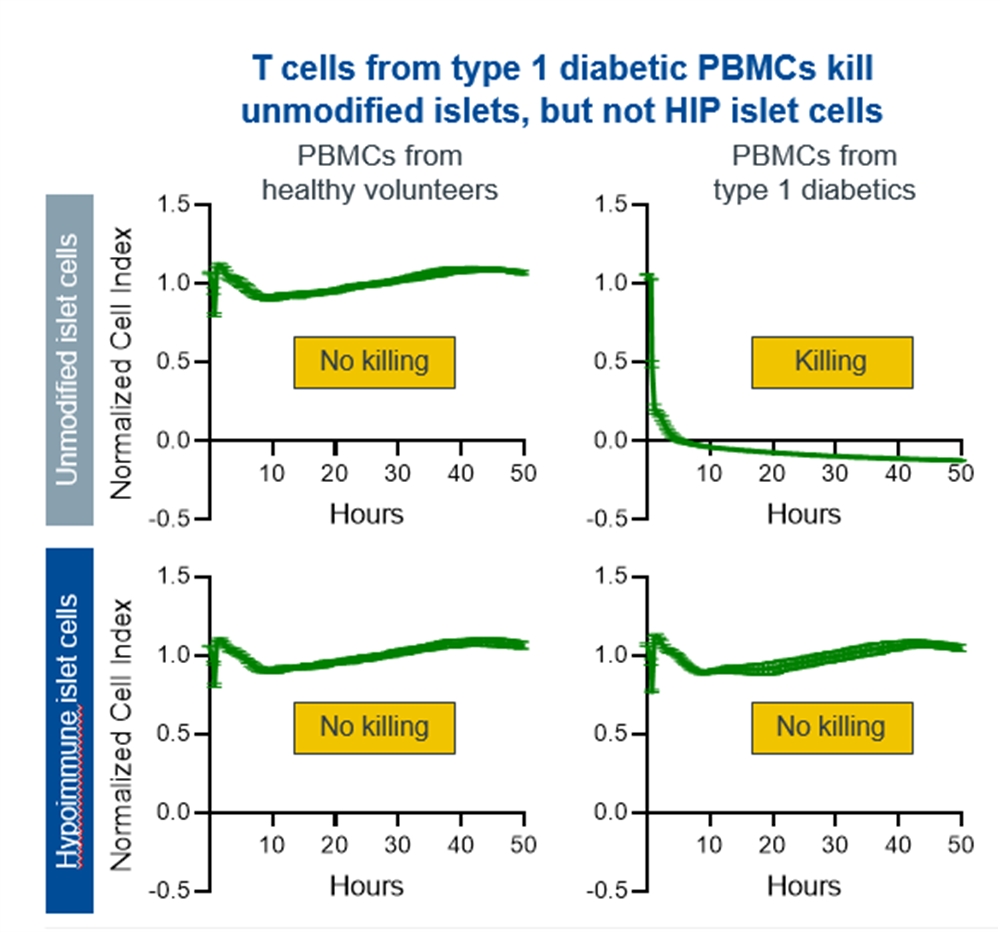
Unmodified human donor derived pancreatic islets (Row 1) are rapidly killed by PBMCs (immune cells) from Type 1 Diabetic donors but are not recognized by immune cells from healthy volunteers. Human donor derived pancreatic islets bearing hypoimmune modifications (Row 2) are not recognized by the PBMCs of healthy volunteers or type 1 diabetics
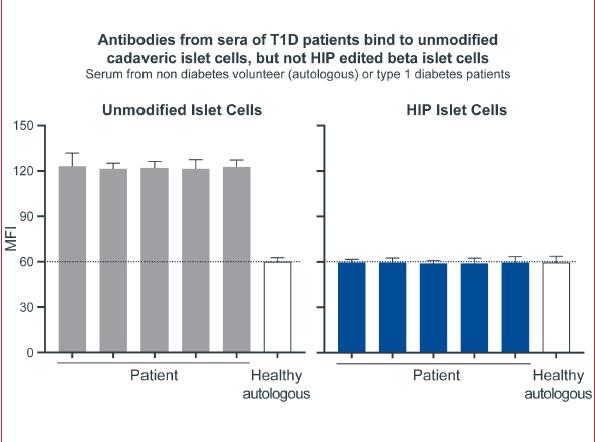
Left graph: Unmodified human donor derived pancreatic islets (unmodified islet cells) are readily recognized by antibodies in the serum of Type 1 Diabetic patients. Antibody binding is corelated with higher values of Maximum Fluorescent intensity (MFI) Right graph: Human donor derived pancreatic islets bearing hypoimmune modifications are not recognized by antibodies in serum of healthy volunteers or Type 1 diabetic patients
27
Hypoimmune human donor pancreatic islet cells survive in humanized mice and regulate glucose levels
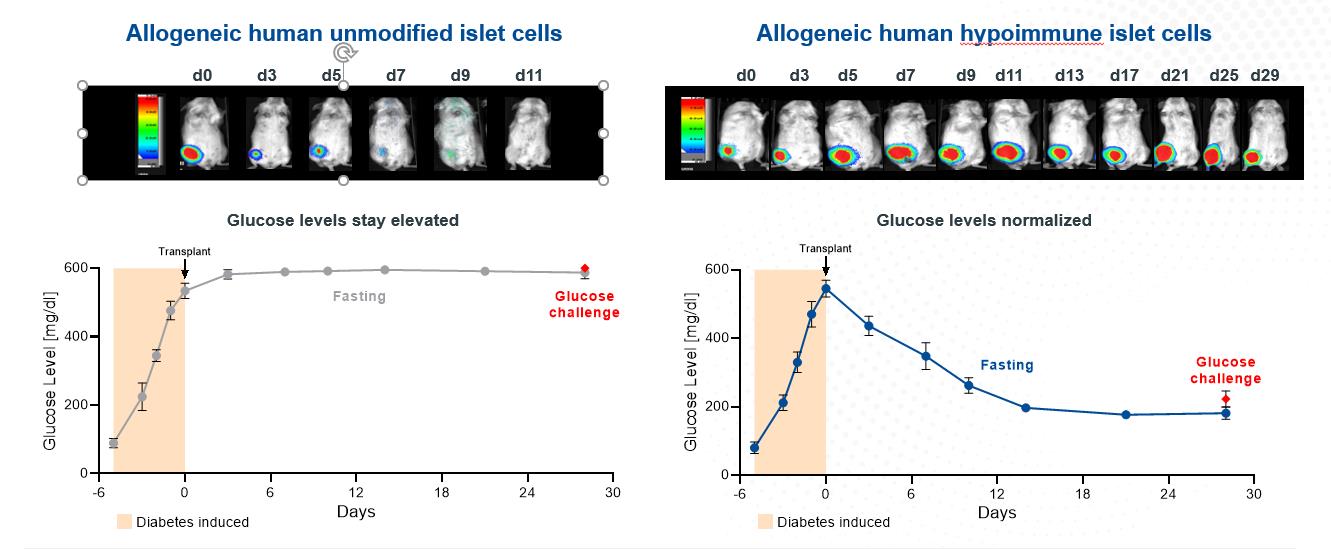
Top image panel on left: Unmodified human donor derived pancreatic islets injected intramuscularly into humanized mice are rejected by recipient mice by D9. Viable cells are marked via a bioluminescent marker that is lost when transplanted cells are no longer viable. Top image panel on right: By contrast hypoimmune human donor derived pancreatic islet cells survive in humanized recipient mice till D29 when the experiment was terminated
Lower graphs are serum glucose measurements in the humanized mice. Diabetes was induced via streptozotocin treatment that results in loss of glycemic control as visualized by an immediate and sustained increase in serum glucose. Transplantation of unmodified human donor derived pancreatic islets had no effect on serum glucose levels. By contrast hypoimmune human donor derived islets successfully lowered serum glucose levels as measured by a glucose tolerance test.
Development Plan and Key Next Steps
Our work is currently focused on manufacturing GMP-grade, gene-edited, pluripotent stem cell banks; scaling manufacturing; and characterizing the product. We are working through the process development and IND-enabling studies to allow for an IND submission for SC451 as early as 2023.
GPC Program
Our GPC program aims to deliver healthy allogeneic GPCs, the precursors to both astroglia and myelin-producing oligodendrocytes. This program has the potential to treat myelin and glial-based disorders, which represent a broad group of debilitating neurological disorders, such as multiple sclerosis and a number of neurodegenerative disorders, none of which have effective treatment alternatives. We intend to develop our stem cell derived GPC therapies for secondary progressive multiple sclerosis, Pelizaeus-Merzbacher disease other disorders of myelin, Huntington’s disease, and other astrocytic diseases. Our goal is to submit three INDs for SC379 in the next several years.
Background on Myelin and Glial Based Disorders
Glial cells are the support cells of the human CNS. The two major types of CNS-derived glial cells are oligodendrocytes—the cells that produce myelin, the insulating substance of the brain’s white matter that enables neural conduction and astrocytes, the support cells of neurons and their synapses. These two kinds of glial cells arise from human GPCs (hGPCs), are responsible for remyelination in the injured and demyelinated adult brain and spinal cord.
Diseases of glial cells are among the most prevalent and disabling conditions in neurology. These disorders include the disorders of oligodendrocyte loss and myelin failure and the disorders of astrocytes, which include a number of neurodegenerative and psychiatric disorders. What all these disorders have in common is a significant glial contribution to their pathogenesis, and a lack of disease-modifying treatment options.
28
Congenital Leukodystrophies. A number of hereditary disorders of oligodendrocyte loss or dysfunction are characterized by a failure in myelin synthesis or structural stability. Tens of thousands of children in the United States suffer from diseases of myelin loss. The most prototypic example of this class of diseases is Pelizaeus-Merzbacher disease (PMD), an X-linked leukodystrophy most often manifesting in male infants and young boys, caused by mutations in the oligodendrocytic PLP1 gene, which results in widespread hypomyelination. There is no treatment for PMD, which is typically fatal in childhood. We intend to deliver intracerebral transplants of stem cell-derived GPCs to the brains of PMD patients, with the goal of replacing PLP1 mutant oligodendrocytes with healthy cells capable of producing normally compact myelin. Prevalence of PMD in the general population is estimated to be approximately 1 in 100,000 in the United States. While we are initially targeting PMD as our proof of concept, congenital leukodystrophies as a group affect a more significant population, or about 1 in 7,600 births.
Multiple Sclerosis (MS). MS is a debilitating disease characterized by both inflammatory myelinolysis and degenerative axonal loss. There are two major forms, the initial relapsing remitting form, known as RRMS, and its later progressive neurodegenerative phase designated secondary progressive MS (SPMS). RRMS is characterized by clearly defined attacks with new or increasing neurologic symptoms. In contrast, SPMS is characterized by progressive neurodegeneration with a loss of neurons, including those that were previously demyelinated during the RRMS phase of the disease. The demyelination occurs in a diffuse fashion throughout the adult brain and appears to reflect a loss of axonal support by local oligodendrocytes. The delivery of GPCs into such chronically demyelinated brain may offer tangible benefits through the oligodendrocytic engagement of axons, as well as by myelin repair. MS is highly prevalent, with estimates of up to 1.0 million in the United States, 600,000 in Europe, and 2.8 million patients globally. Approximately 85% of MS patients receive a diagnosis of RRMS initially while 15% of patients are diagnosed with primary progressive MS (PPMS). Up to a third of RRMS patients transition to secondary progressive MS within a decade if untreated, and most will progress to SPMS within 20-25 years of diagnosis. Success with a stem cell derived GPC product in SPMS, and especially with a hypoimmune product, could enable further expansion into the RRMS patient population.
Huntington’s Disease (HD). HD is a neurodegenerative disorder in which glial pathology appears to make a significant causal contribution. It is an autosomal dominant disorder characterized by abnormally long CAG repeat expansions in the first exon of the Huntingtin gene. The encoded polyglutamine expansions of mutant huntingtin protein disrupts its normal functions and protein-protein interactions, ultimately yielding widespread neuropathology, most rapidly evident in the neostriatum. We have found that glial pathology is a major contributor to the functional deficits of HD and repairing the glial pathology has significant and positive effects in animal models. There are approximately 41,000 symptomatic Americans and more than 200,000 at-risk of inheriting HD. In Europe, there are approximately 50,000 patients with HD.
Current Treatment Landscape and Unmet Need
Congenital Leukodystrophies. There are no viable treatment options for these conditions, only supportive and palliative therapies for symptoms as they present.
MS. Current treatments for MS are largely limited to treatments for RRMS; few treatments are approved for SPMS, and these have at best marginal efficacy in delaying disease progression; none are restorative. Currently approved treatments for RRMS may be divided into three broad categories of disease modifying therapies: (i) first line injectables (such as beta-interferons, Copaxone), (ii) newer oral agents (such as Tecfidera, Gilenya, Mayzent, Zeposia), and (iii) high-efficacy agents (such as Tysabri, Lemtrada, Ocrevus). Despite many recently successful drug launches in the RRMS space, these drugs still only slow the progression of disease and aid in the recovery from attacks, and there remains no treatment that confers functional restoration or effective cure for this disease.
HD. Currently, there is no treatment to stop or reverse Huntington’s disease. Treatment is limited to several medications that can help minimize symptoms, including the drug tetrabenazine, antipsychotic drugs, antidepressants, and tranquilizers.
Our GPC Program Approach
Our approach to treat myelin and neurodegenerative disorders is via the delivery of healthy allogeneic stem cell-derived GPCs. We have developed methods for producing and isolating these cells from pluripotent stem cells and delivering them in the purity and quantities necessary for their replacement of endogenous diseased cells. We believe that both the myelin disorders and glial-based neurodegenerative conditions have compelling potential for our ex vivo therapy.
Preclinical Data
Congenital Leukodystrophies. The capacity of stem cell-derived GPCs for remyelination has been conducted in animal models of congenital hypomyelination. Our collaborators used newborn shiverer mice that have a genetic defect in myelin basic protein (MBP), resulting in their neurons being hypomyelinated and the mice having a shortened lifespan. When iPSC-derived hGPCs were transplanted into these mice, the cells spread widely throughout the brain, developing as astrocytes and oligodendrocytes. These oligodendrocytes generated mature myelin that effectively restored neuronal conductance and prolonged survival in the transplanted
29
mice. We believe that these data suggest the feasibility of iPSC-derived hGPC implantation in treating childhood disorders of myelin formation and maintenance, as depicted in the figure below:
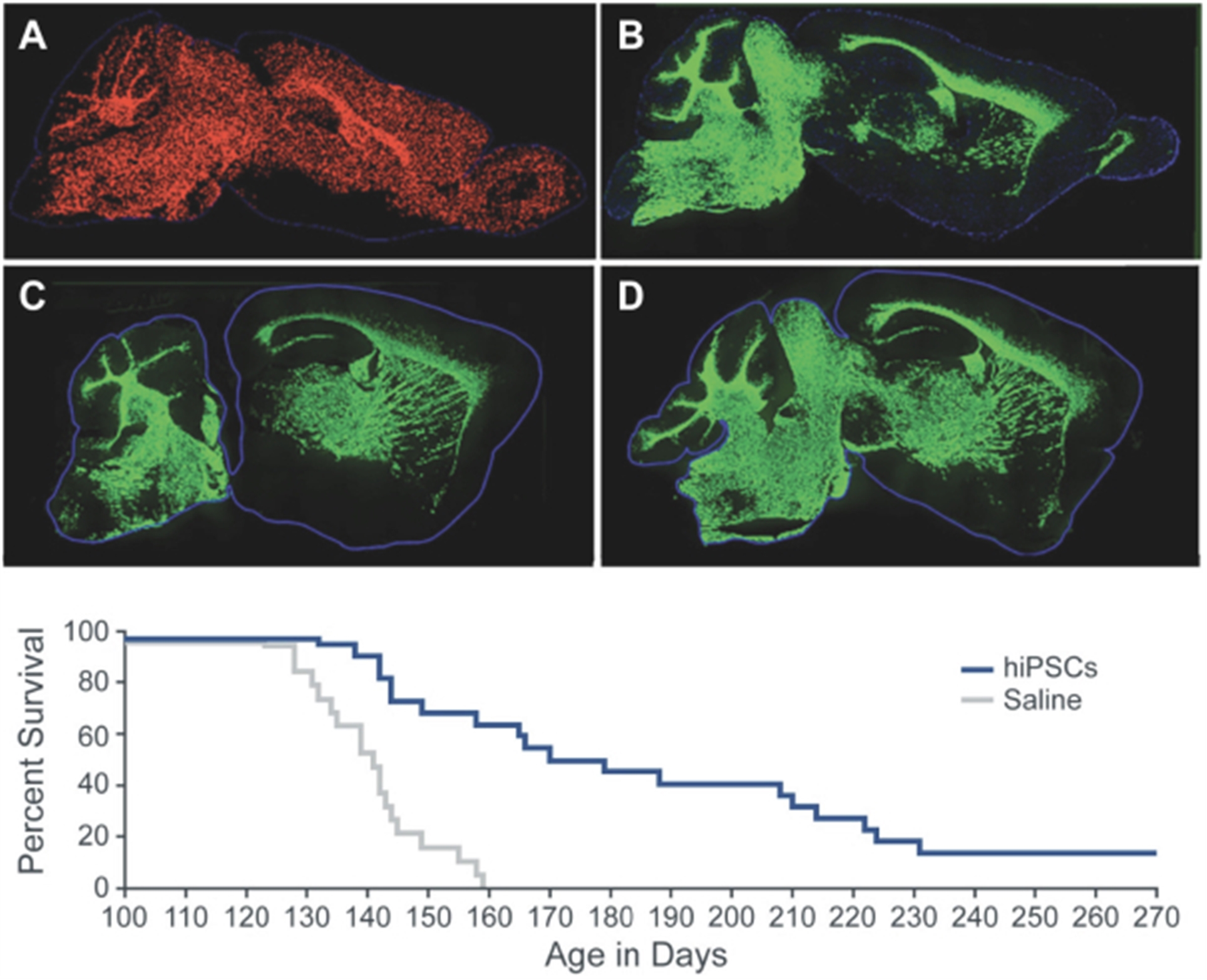
hGPCs myelinate widely to greatly extend the survival of hypomyelinated mice. A, Dot map indicating distribution of human iPSC-derived GPCs at 7 months of age, following neonatal engraftment in a shiverer mouse brain. Widespread colonization and chimerization of the host brains by iPSC-derived hGPCs is evident (human nuclear antigen, red). B, iPSC-derived hGPC-derived myelination in shiverer forebrain, at 7 months; section 1 mm lateral to A. Myelin basic protein (MBP)-immunoreactivity (green) is all human donor-derived. C, D. Myelination in sagittal sections taken at different mediolateral levels from 2 additional 7 month-old mice, each engrafted with iPSC-derived hGPCs at birth. E, Kaplan-Meier plot of survival of iPSC-OPC implanted (n=22) vs. saline-injected (n=19) control mice. Scale: A-B, 2 mm. Adapted from Wang, Cell SC 2013.
MS. Our prior studies established the ability of stem-cell derived hGPCs to myelinate the developing shiverer brain and rescue the afflicted mice; however, the experimental subjects were neonates, not adults. Until recently, it was unclear whether GPCs are able to migrate extensively in adult brain tissue, as would be required for the repair of diffusely demyelinated adult brains. To explore whether the introduction of stem-cell derived hGPCs delivered directly into the adult brain could remyelinate axons in the setting as might be encountered clinically in multiple sclerosis our collaborators studied three different biologic models. First, it was shown that stem-cell derived hGPCs can disperse within and myelinate the brains of adult shiverer mice (as depicted in the figure below). Second, it was shown that neonatally-engrafted hGPCs engrafted as a neonate can generate new oligodendrocytes and remyelinate demyelinated axons after chemically induced demyelination. This result demonstrated the ability of already-resident hGPCs to remyelinate previously myelinated axons after a new demyelinating insult as an adult, as well as the ability of transplanted hGPCs to reside as a functional reservoir of new myelinogenic cells in the host brains. Third, it was shown that hGPCs transplanted into the adult brain after chemically induced demyelination can remyelinate denuded axons. These data suggest that transplanted hGPCs can disperse broadly and differentiate as myelinogenic cells in the adult brain, and that they are able to remyelinate demyelinated axons and white matter lesions of the brain after an insult as an adult.
30
hGPCs Mediate Robust Myelination After Transplantation into the Adult Shiverer Brain
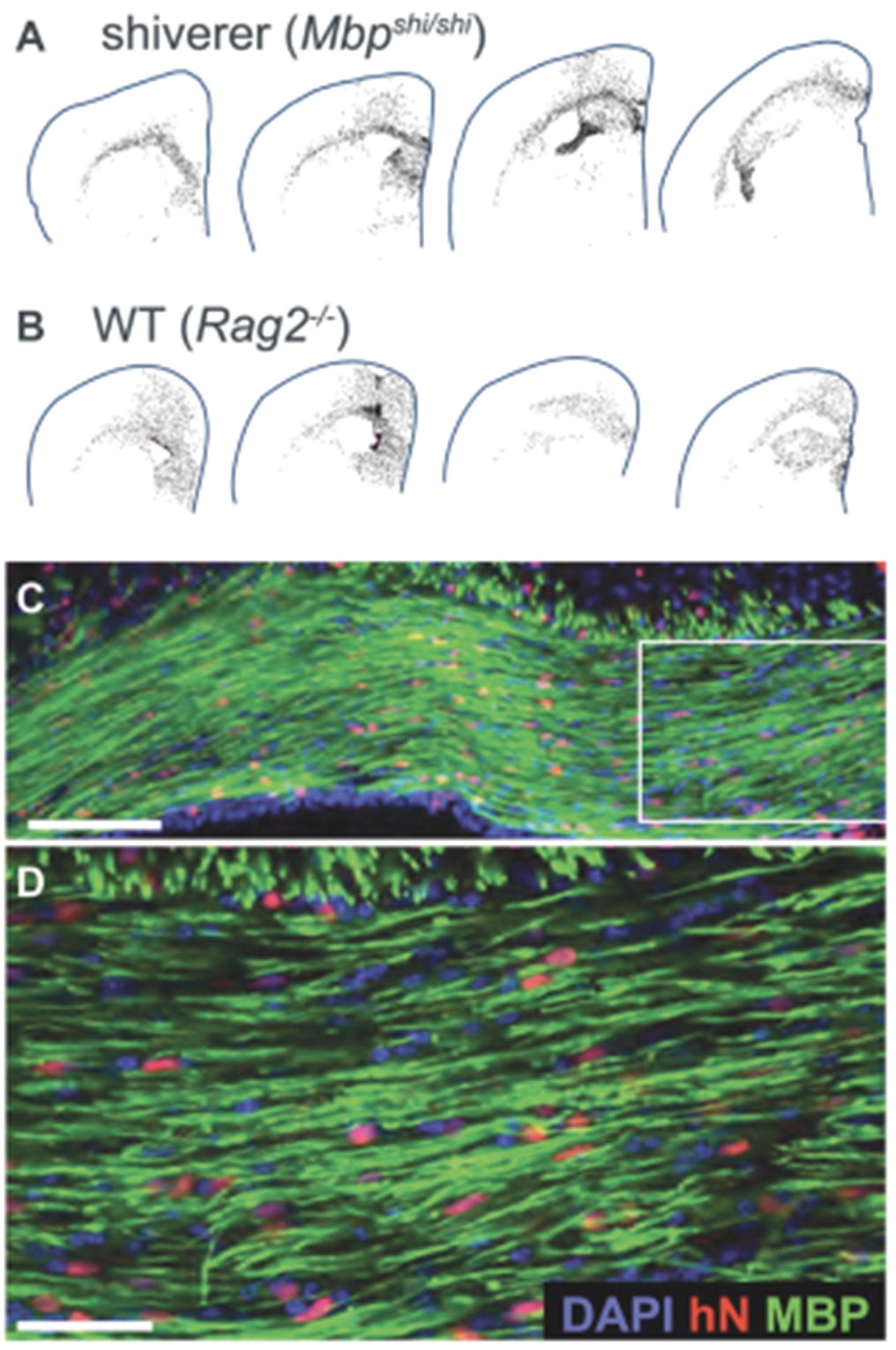
Human GPCs proved both highly migratory and robustly myelinogenic, after delivery to the hypomyelinated adult shiverer x rag2-/- brain. A, By 19-20 weeks of age (mice were injected as post-weaning adults, at 4-6 wks) the injected cells had dispersed broadly throughout the forebrain white matter. B, hGPCs delivered to myelin wild-type rag2-/- mice distributed throughout both gray and white matter. C, Oligodendrocyte differentiation and myelinogenesis by donor hGPCs was robust, with myelination of brain regions that would typically be demyelinated in shiverer mice D, a higher power image of C shows the high proportion of donor cells in those brain regions. Note that DAPI marks all nuclei, hN marks the hGPCs, and MBP marks the remyelinated regions in C and D. From Windrem et al, Cell Reports 2020.
HD. Our collaborators explored the cellular basis for HD related glial pathology and identified significant defects in potassium channel and glutamate uptake mechanisms in HD glia, which appeared to account for both the glial pathology and its deleterious effects on synaptic function. Together, these studies suggest a critical role for glial pathology in the progression of HD and suggested the potential for glial cell replacement as a therapeutic strategy in HD, and more broadly, to other neurodegenerative diseases in which glial pathology might be causally contributory. It was confirmed in preclinical mouse studies that stem-cell derived hGPC transplant ameliorated both the neuronal and glial pathology of HD by restoring synaptic homeostasis and normal synaptic function to the most affected regions of the host brain.
The majority of the studies with human GPCs thus far have been xenogeneic grafts of human GPCs to neonatal or adult mice or rats (and in a small sample POC study limited to adult tissue-derived hGPCs, NHPs). Our collaborators have also performed studies with murine GPCs transplanted into both developing and adult mice, which have confirmed allogeneic GPC migration and integration. However, we have no assurance that human GPC engraftment of human brain will result in the widespread migration and colonization of host brain that is seen with xenogeneic grafts. To better model the human-to-human graft paradigm, our collaborators have established a new model to evaluate if GPC engraftment will result in migration and colonization in a host brain. This model allows observation of the competitive interactions of the two separately tagged human GPC populations. The human-into-human grafts expanded and integrated well in their humanized host, with competitive interactions. As might be anticipated in the clinical setting of healthy cells being transplanted for the purpose of replacing lost or diseased hGPCs, the healthy donor cells outcompete both diseased and older cells to ultimately colonize the hosts. These data have provided preclinical assurance of the fundamental premise of our approach, that healthy human donor cells can replace lost or diseased human cells in vivo. That said, this determination remains to be made in patients.
31
GMP Grade Stem Cell Derived hGPCs for Clinical Studies
A protocol to direct differentiation of human ESCs, as well as iPSCs, to hGPCs has been established. These hGPCs cells remain bipotential for astrocytes and oligodendrocytes, and they differentiate to either fate depending on local signaling.
This protocol has been transferred to a GMP facility in order to enable production of clinical grade cells for both safety and efficacy testing. These cells have been validated to robustly remyelinate shiverer mouse brains upon intracerebral transplantation. We plan to use these cells for our IND-enabling studies and initial clinical trial material.
Development Plan and Key Next Steps
Progression of SC379 to IND is planned to follow completion of definitive safety and toxicology studies. Definitive preclinical efficacy studies using the anticipated clinical product are also planned and will replicate studies that we have published. Since GPCs are not a terminally differentiated cell type and divide and differentiate in vivo post-transplantation, we will continue to assess potential safety risks, including the risk of tumorigenicity. We expect to submit IND applications for SC379 for SPMS, PMD, and HD in the next several years.
Cardiomyocyte Program
Background on Heart Failure
Heart failure (HF) is a classic example of a disease of cell loss, ideally suited to the application of ex vivo engineered cells. The clear but ambitious goal of our program is to replace missing cells after a myocardial infarction, commonly known as a heart attack, in an attempt to restore heart function and improve outcomes for patients. HF is a life-threatening syndrome, and patients with HF have a mortality rate of 20-30% within one year of diagnosis and a mortality rate of around 50% within five years of diagnosis.
HF with reduced ejection fraction (HFrEF), is a severe form of HF where heart muscle is unable to contract, and therefore pump, adequately. HFrEF is most frequently a consequence of a loss of heart muscle cells (cardiomyocytes), following a myocardial infarction. In the United States, there were approximately 380,000 deaths associated with HF in 2018 according to the United States Centers for Disease Control and an overall prevalence of approximately 6 million people with HF, with similar numbers in Europe.
In general, HF has been a challenging area for drug and device development, including only one new drug, Entresto, approved in the last 20 years, and a limited number of devices introduced including electrical resynchronization therapy and implantation of left ventricular assistance devices (LVADs). These approaches provide only symptomatic relief and do not address the underlying loss of cardiomyocytes associated with HFrEF. As a result, HFrEF currently remains a progressive and deadly disease with a large unmet need worldwide.
To date, efforts to develop cell-based therapies to address this unmet need have provided little evidence of clinical benefit. Importantly, these attempts have typically utilized cells such as bone marrow-derived mononuclear cells and mesenchymal stromal cells where any potential benefit would be limited to paracrine mechanisms and not the direct replacement of lost cardiomyocytes.
Our cardiomyocyte program aims to directly regenerate the heart, by replacing lost cardiomyocytes with iPSC-derived cardiomyocytes, with the goal of restoring heart muscle and increasing ejection fraction, which is the percentage of blood the heart pumps with each heartbeat. Replacement of lost cardiomyocytes with iPSC-derived cardiomyocytes that engraft and function correctly has the potential to prevent or even reverse the progression of HFrEF.
Developing an ideal stem cell-derived cardiomyocyte therapy involves many steps, including:
|
|
• |
differentiating cardiomyocytes at scale that engraft upon transplantation, beat in synchrony with the host heart muscle, and improve heart function; |
|
|
• |
engineering cardiomyocytes to avoid rejection due to the host immune response to the transplanted cells, without requiring immunosuppression; and |
|
|
• |
addressing the risks associated with potential transient arrhythmias, or temporary abnormal heart beats, following transplantation. |
Differentiating Cardiomyocytes at Scale that Engraft, Beat Correctly and in Synchrony with the Host Heart Muscle, and Improve Host Heart Function
Scientists, including Dr. Chuck Murry, our Senior Vice President, Head of Cardiometabolic Cell Therapy, have been working for over 20 years towards developing a regenerative therapy for HFrEF with the goal of transplanting cardiomyocytes derived from
32
human pluripotent stem cells that engraft, function, and persist in the human heart in vivo. The groundwork for potential future clinical development has been laid by key breakthroughs such as the ability to direct the differentiation of stem cells selectively into cardiomyocytes, including producing pharmaceutical grade cardiomyocytes at large scale in bioreactors, and the ability to transplant such cardiomyocytes to induce remuscularization of injured hearts.
Initial preclinical attempts to remuscularize the infarcted heart were unsuccessful due to death of the transplanted cells within a few days of delivery. None of the animals with failed engraftment showed improvement in cardiac function, indicating that engraftment is essential for functional improvement. Our collaborators developed a pro-survival cocktail that kept cells alive through the rigors of transplantation, allowing the cardiomyocytes to self-assemble into new muscle tissue and induce ingrowth of new blood vessels and connective tissue from the surrounding heart muscle. Once engraftment was successful, cardiac function improved. As our collaborators’ capabilities to scale cell manufacturing increased, studies progressed from mice to rats to guinea pigs, all showing improved function:
Human ESC-Cardiomyocytes Improve Function in Injured Rat and Guinea Pig Hearts
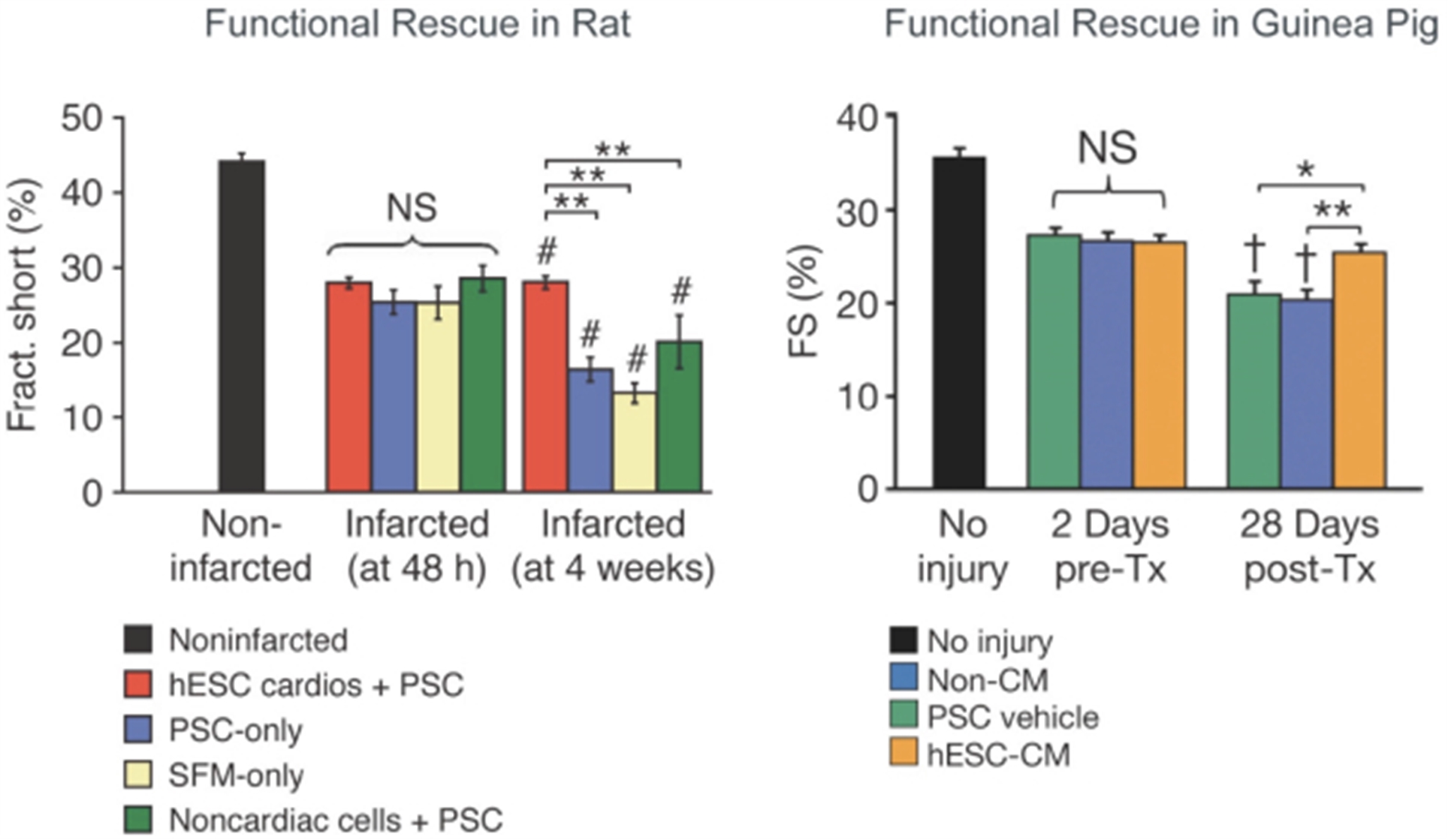
Left panel: functional rescue in rat. All groups showed comparably reduced fractional shortening after infarction at 2 days pre-transplantation (Pre-Tx). At 28 days post-transplantation there was preservation of fractional shortening in animals receiving hESC-cardiomyocytes, with deterioration of function in all other groups. PSC, pro-survival cocktail. SFM, serum-free media. **p<0.01; # p<0.05 vs. 2 days Pre-Tx. Right panel: functional rescue in guinea pig. Following cardiac injury at 2 days before transplantation, all groups showed comparably reduced fractional shortening. At 28 days post-transplantation there was preservation of function in animals receiving hESC-cardiomyocytes (hESC-CM), with deterioration of function in other groups. *p<0.05. **p<0.01. †p<0.05 vs 2 days Pre-Tx. From Laflamme et al, Nature Biotechnology 2007 (left) and Shiba et al, Nature 2012 (right).
Current methods demonstrate regeneration of the hearts of large animals including pigs and NHPs by transplanting human ESC-derived cardiomyocytes (hESC-CM). The figure below shows low magnification microscopic images from NHP hearts that were infarcted and then received either hESC-CM or saline controls. The replacement of heart muscle by scar tissue is evident in the saline-treated heart, whereas human heart muscle has repopulated the infarct in the hESC-CM treated group.
33
Remuscularization of the Heart of an NHP by hESC-CM Transplantation
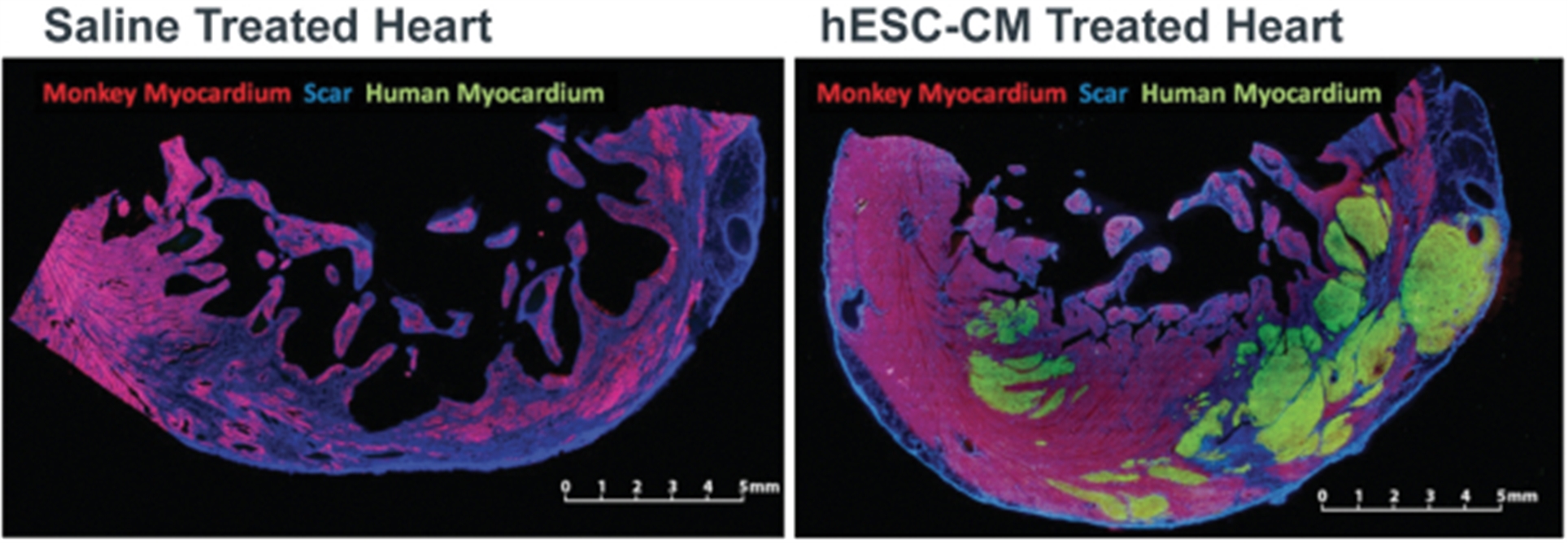
The saline-treated heart (left) shows infarct scar tissue (blue) replacing the myocardium lost to infarction. The hESC-CM treated heart (right) shows a large graft of human heart muscle (green) replacing the myocardium lost to infarction. From Liu et al, Nature Biotechnology 2018. Scale bar, 5 mm.
We conducted an experiment to help us understand the mechanism of action and learn whether the transplanted muscle beat in synchrony with the host heart muscle. hESC-CMs were genome-edited to express a protein that fluoresced green with each contraction, and their behavior was studied after transplantation into an infarcted macaque heart. hESC-CMs showed 1:1 synchrony with the host heart, indicating that the graft follows the heart’s natural pacemaker, an essential result for heart regeneration.
A final question was whether this regeneration improves the function of the injured heart. To assess this, engrafted NHP hearts were studied by magnetic resonance imaging (MRI), the gold standard for assessing cardiac contractile function. As illustrated in the figure below, myocardial infarction induced a 25-point drop in left ventricular ejection fraction, the fraction of blood ejected from the heart with each beat.
Control animals receiving a saline injection showed no significant improvement at 4 or 12 weeks, as expected. In contrast, four weeks after receiving hESC-CMs, ejection fraction improved by approximately 10 points, and by 12 weeks, it had improved by a total of approximately 22 points. While the number of animals followed for 12 weeks is limited, cardiac remuscularization in this study restored ventricular function back into the normal range. In contrast, the current standard of care for myocardial infarction, including reperfusion via angioplasty, ACE inhibitors, and beta blockers, increases ejection fraction by approximately 6 points.
34
Restoration of Cardiac Function in NHPs by Transplantation of Human ESC-derived Cardiomyocytes (hESC-CM)
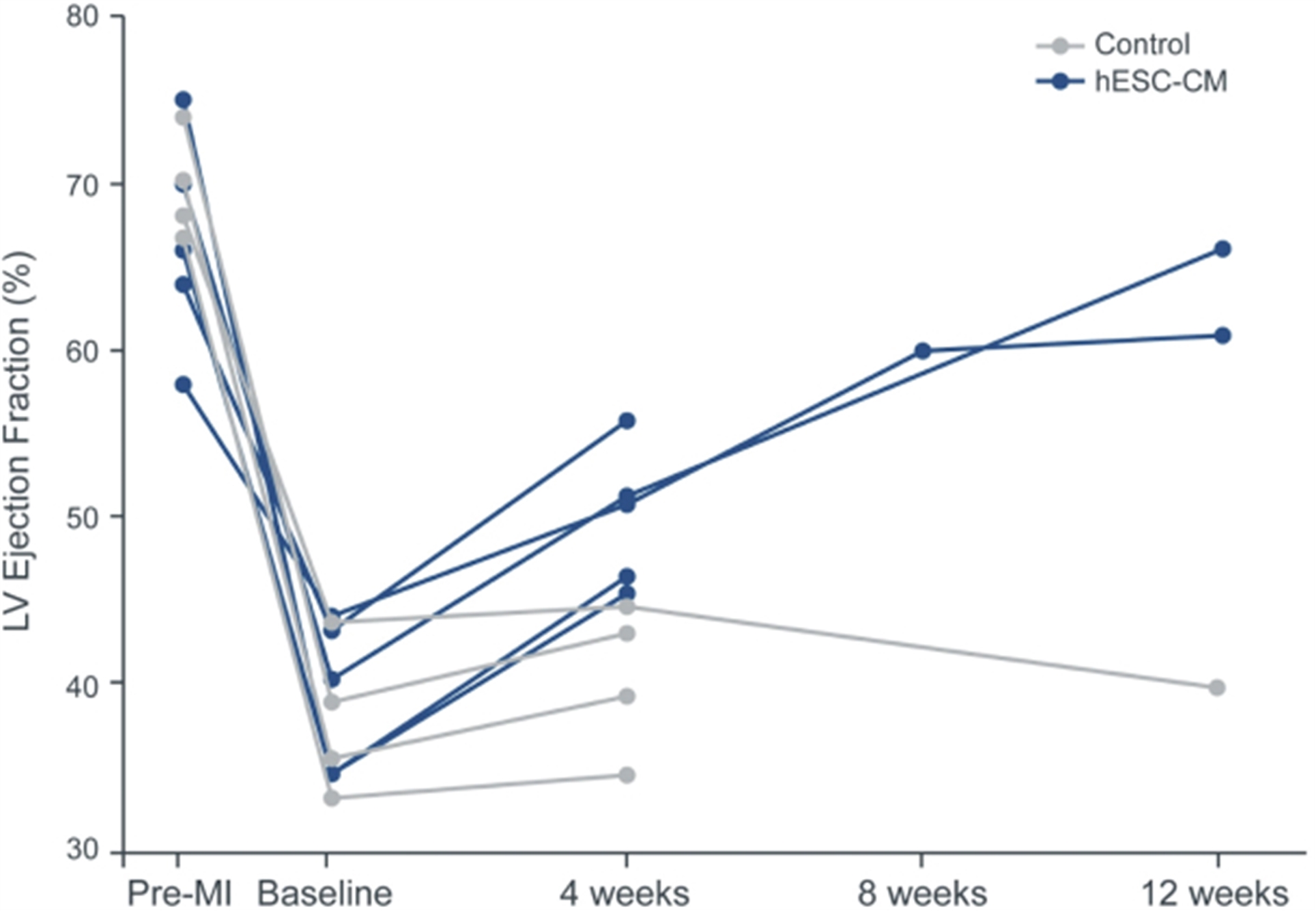
Myocardial infarction reduces ejection fraction (a measure of cardiac function), and there is no spontaneous recovery in control animals receiving a saline injection (gray). All animals receiving hESC-CM (blue) showed significant improvement by 4 weeks, and by 12 weeks after treatment, cardiac function was restored to the normal range.
In summary, structural data demonstrating extensive remuscularization of the infarcted heart in conjunction with physiologic and pharmacodynamic data provide evidence that the transplanted cardiomyocytes directly restore heart contractile function.
Engineering the cells to avoid rejection due to the host immune response to the transplanted cells
Initially, we plan to establish safety with first-in-human clinical trials of our cardiomyocyte cell therapy using immunosuppression to reduce the risk of a host immune response to allogeneic transplanted cells and the potential immune rejection. Our collaborators have studied immunosuppressive regimens in NHP by transplanting rhesus cardiomyocytes derived from stem cells into the hearts of mismatched recipient NHP. An immunosuppressive regimen was identified that keeps the allogeneic grafts alive long term and is considerably less toxic than regimens used for heart transplantation. However, an approach that obviates the need for an immunosuppression regimen has the potential to improve safety and patient eligibility. Therefore, as part of our program lifecycle we intend to switch to a hypoimmune stem cell-derived cardiomyocyte over time, as this should allow us to eliminate or reduce the immune suppression required for durable maintenance of these cells.
Addressing the risks associated with potential transient arrhythmias, or abnormal heart beats, following cell transplantation
The term engraftment arrhythmia refers to a transient period of unstable electrical activity that occurs in some species over approximately four weeks following cardiomyocyte transplantation. Engraftment arrhythmias were not observed in mice, rats, or guinea pigs (probably because their natural heart rates are too fast), but they are observed in NHPs, where they cause mild symptoms, and in farm pigs, where they cause more significant symptoms. The arrhythmias follow a stereotypical course, where they increase in frequency and duration, plateau for a variable period, and then wane until the heart has normal rhythm once again. Once the heart rhythm stabilizes, the arrhythmias seem to disappear permanently. We are exploring three ways to address engraftment arrhythmias: pharmaceutical interventions, genetic modifications to the cardiomyocytes, and adjusting the stage of differentiation of the cardiomyocytes. Electrical mapping studies in NHPs and pigs suggest that the engraftment arrhythmias originate from the site of the cell injection, likely from the injected cardiomyocytes. The stem-cell derived cardiomyocytes in these experiments are more similar to fetal cardiomyocytes than their adult counterparts, and while this favors their engraftment it potentially increases the risk for the
35
induction of arrhythmias. We compared the expression and function of ion channels, proteins regulating the electrical properties of cardiomyocytes, in immature versus mature cardiomyocytes. We identified multiple ion channels which showed differential activity, including HCN4, CACNA1H, and SLC8A1 which are active, as well as KCNJ2 which is inactive, in immature cardiomyocyte relative to mature ones. We subsequently assessed the impact of modulation of a variety of combinations of ion channels in animal studies. We found that PSC-derived cardiomyocytes in which the expression of HCN4, CACNA1H, and SLC8A1 has been abolished and the expression of KCNJ2 has been activated (“MEDUSA cells”) creates cardiomyocytes that lack endogenous pacemaking ability but can follow exogenously supplied electrical stimulation. MEDUSA cells robustly engraft but do not induce arrythmias in a pig model. We are currently testing whether MEDUSA cells can effectively restore heart function in infarcted NHPs as well as the longer-term potential impacts of modifying these ion channels in cardiomyocytes.
MEDUSA cells do not induce arrythmia after transplantation in a pig model
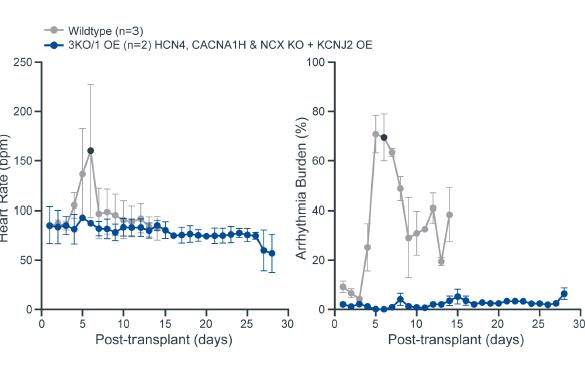
Wildtype stem cell-derived cardiomyocyte grafts induce arrhythmias after transplantation into pig hearts (blue circles), shown by the rise in heart rate and the increased arrhythmia burden (percentage of day spent in arrhythmia). Black circle indicates one wildtype animal that died from complications of the arrhythmia. In contrast, MEDUSA-edited cardiomyocytes (3 KO/1 OE; orange squares) do not induce arrhythmias.
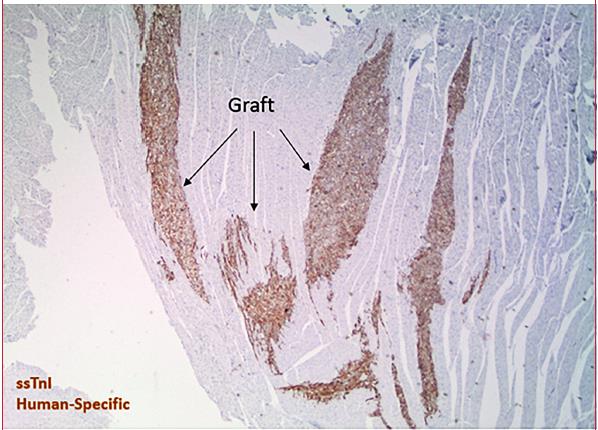
Immunostaining for human cardiac muscle (human-specific slow skeletal troponin I stain, brown) demonstrates large grafts of human myocardium in the heart of a pig receiving MEDUSA cells. Thus, absence of engraftment arrhythmia is not due to absence of engraftment.
36
Development Plan and Key Next Steps
Our key milestones include understanding any potential impact on function and safety of these MEDUSA edits, their ability to protect longer-term from the risk of arrhythmias, completing GLP toxicology studies, and additional efficacy and safety studies in NHPs and pigs, with the goal of submitting an IND in the next several years. We plan to finalize our clinical plan based on the results of ongoing studies in NHPs using the MEDUSA edits in stem-cell derived cardiomyocytes as well as early results from humans in other settings with our hypoimmune platforms.
Our in vivo Cell Engineering Platform
Overview
In vivo cell engineering aims to treat human disease by delivering a therapeutic payload to cells inside a patient’s body to repair or control genes. Historically there have been four key challenges to in vivo cell engineering:
|
|
• |
Delivering any payload (such as DNA, RNA, proteins, organelles, integrating versus non-integrating, size), |
|
|
• |
to any cell (by increasing the volume of distribution), |
|
|
• |
in a specific (for instance just T cells), and |
|
|
• |
repeatable way (such as achieving limited immunogenicity to allow re-dosing). |
Our in vivo cell engineering platform is focused on engineering fusogens that, when combined with delivery vehicles, can effectively deliver a payload to a desired cell or location in the appropriate quantities in vivo. The combination of a fusogen with a delivery vehicle referred to as a fusosome. We believe our platform provides us with the flexibility to deliver a wide range of payloads to make different modifications for different diseases, as well as delivery vehicle options to address volume of distribution and re-dosing, which could fundamentally expand the treatment potential of in vivo therapies.
Our Approach to Building our in vivo Cell Engineering Platform
We have approached the development of our in vivo cell engineering platform by investing in solutions to overcome the key challenges outlined above:
|
|
• |
Delivery. We believe the critical limitation for in vivo cell engineering is delivery, and therefore, we are investing significantly in delivery technologies, including our fusogen technology, which is designed to enable both cell-specific delivery and delivery of diverse payloads. We were founded with core technology in this area which was the product of a multi-year effort by a Flagship Labs innovation team at Flagship Pioneering led by Dr. Geoffrey von Maltzahn, one of our board members. This effort is led by Dr. Jagesh Shah, our VP, Gene Therapy Technologies. |
|
|
• |
Gene modification. There has been substantial recent progress in gene modification and the field is now at the point where virtually any desired modification can be performed in vitro. However, no single technology or platform is optimal for all possible applications. To this end, we are developing capabilities across multiple technologies and investing to develop our own novel technologies to be applied on a case-by-case basis, an effort that is led by Dr. Ed Rebar, our Senior Vice President, Chief Technology Officer. We also have entered, and intend to enter more, agreements with other companies that have capabilities in this area. |
|
|
• |
Manufacturing. We are investing proactively in process development, analytical development, CMC regulatory, supply chain, quality, and other manufacturing sciences in order to enable scalable manufacturing of our in vivo therapies and ensure broad access. We have also built a pilot manufacturing plant in South San Francisco, California and entered into a long-term lease agreement for a facility in Fremont, California, where we intend to build our own clinical trial and commercial GMP manufacturing capabilities. These efforts are led by Dr. Stacey Ma, our Executive Vice President, Technical Operations. |
Our Approach to Building our in vivo Cell Engineering Portfolio
We have prioritized cell types for our programs where:
|
|
• |
existing proof of concept in humans and animal models demonstrates that in vivo cell engineering should have a clinical benefit; |
|
|
• |
high unmet need can be addressed by modifying a particular cell type; |
|
|
• |
delivery is the most critical bottleneck, such that delivering payloads specifically to the target cell type could lead to highly differentiated and transformative therapeutics; and |
37
|
|
• |
an opportunity to apply the technology more broadly exists, which creates the potential for more medicines if successful (for example, delivery to hepatocytes unlocks potential to treat many diseases with different payloads). |
Based on this prioritization, we are initially focused on three cell types: T Cells, Hepatocytes, Hematopoietic Stem Cells
History of in vivo Cell Engineering and Current Limitations
Starting several decades ago, the nascent field of gene therapy focused on experimenting with different means of transmitting genetic payloads via viral vectors. Seminal work by Dr. Richard Mulligan, our Executive Vice-Chairman and Head of SanaX, and colleagues established the promise of gene therapy by delivering genes into host chromosomes, thereby correcting genetic deficits. More recently, significant investments have resulted in improved safety and efficacy of viral vectors. However, most approaches continue to concentrate on adapting the innate capabilities of various viruses to transmit these payloads.
Profound benefits have been realized in cases where there is direct correlation between the biological activity transmitted by the therapy and the genetic activity that is missing in the patient. AAV vectors, which are broadly used by gene therapy researchers due to their broad tissue tropism, lack of pathogenicity, and ability to target both dividing and non-dividing cells. While these therapies have had a categorical impact on their target patient populations, they have only scratched the surface of the potential of in vivo cell engineering, with success limited to a small number of patients.
Broad impact of gene therapies has been limited by challenges within three key areas:
Payload delivery is limited by:
|
|
• |
Limited Cell Specificity. Most commonly used AAV vectors have broad tissue specificities. If a specific type of cell needs to be targeted within a tissue or organ to achieve the desired therapeutic effect, a lack of targeting specificity can result in a limited amount of payload reaching the desired cell. Moreover, the transduction of non-target cells can necessitate the use of high doses of vector to achieve the maximal therapeutic effect in the desired target tissue, which in turn can lead to toxicities due to the transduction of non-target cells, as well as create challenges in manufacturing at adequate scale. Lipid nanoparticles (LNPs) target any cell expressing the LDL receptor, making them both non-specific and mainly absorbed by hepatocytes in the liver when dosed systemically. |
|
|
• |
Limited Volume of Distribution. Volume of distribution refers to the ability of a therapeutic to reach various tissues. While AAV vectors can be used to systemically deliver payloads to certain tissues, such as muscle, in the case of other therapeutically important targets, such as cells of the CNS, only a small proportion of cells can be transduced. |
|
|
• |
Immunogenicity. Most viruses used as vectors elicit an immune response in the patient, causing the patient’s immune system to attack the vector. Previous exposure to the virus used as a vector increases the immune response and may limit the benefit or create safety issues for the patient. Many patients, for example, demonstrate pre-existing antibodies to specific AAV serotypes which can limit transduction efficiencies, and therefore clinical benefit. Furthermore, once an AAV vector is administered to a patient, in most cases the infection leads to an immune response that precludes the ability to re-dose. |
Genome modification is limited by:
|
|
• |
Payload Size and Type Restrictions. The natural genome size of a virus vector imposes a discrete limit on the amount of biological information that can be transmitted. Currently, there exist a number of important disease targets that require the delivery of payloads too large for AAV, which has a maximum payload capacity between 4.5-5kb. In addition to the need to deliver sequences encoding a desired protein that may not fit into an AAV vector, the increasing interest in the use of gene-editing machinery to correct specific gene defect via homologous recombination or transposition will require delivery vehicles capable of a larger payload capacity than is currently available. For most viruses currently used for in vivo therapy, the payload type is generally limited to the specific genetic material of the virus (e.g., DNA or RNA). The ability to deliver additional payloads, such as proteins, could unlock novel therapeutic opportunities. Non-viral delivery with LNPs has been limited to RNA and proteins to date, with an inability to deliver DNA. |
|
|
• |
Durability Limitations. Obtaining the persistence of the desired level of expression over long periods of times can be problematic, due to both immune reactions and the silencing of vector expression. In cases where the target cells are undergoing replication, as can be the case in pediatric patients for example, durability of expression by non-integrating vectors or delivery of material that does not permanently change the cell’s DNA can also be limited by the gradual loss of vector sequences as infected cells replicate. |
38
Execution in manufacturing is limited by:
|
|
• |
Complex manufacturing. Today, the adage of “the process is the product” applies with particular relevance to these in vivo viral vector-based therapies. These therapies are relatively more complex to characterize and control during manufacturing compared to other common biologically derived modalities such as recombinant proteins and antibodies. Similarly, process and analytical sciences that can enable significant scale-up for in vivo therapies are still well behind that of proteins and antibodies. Current vector manufacturing has limited scale and yield, which limits access for patients. |
Our Solution – Fusogen Technology
To address some of the existing challenges of in vivo cell engineering, we are developing our fusogen technology by engineering proteins found in nature to enable the delivery of any payload to specific cells.
Background on Fusogens
Fusogens are a well-studied class of naturally occurring proteins that mediate the trillions of cell-to-cell and intracellular fusion events occurring in the human body every second. In 2013, the Nobel Prize in Physiology or Medicine was awarded for the elucidation of the roles of fusogens in mediating intracellular trafficking in nature. First, fusogens enable recognition of a specific target membrane. Second, they promote membrane fusion by acting as thermodynamic engines for opposing membranes, pulling them together and thereby promoting fusion.
39
Our Fusogen Technology
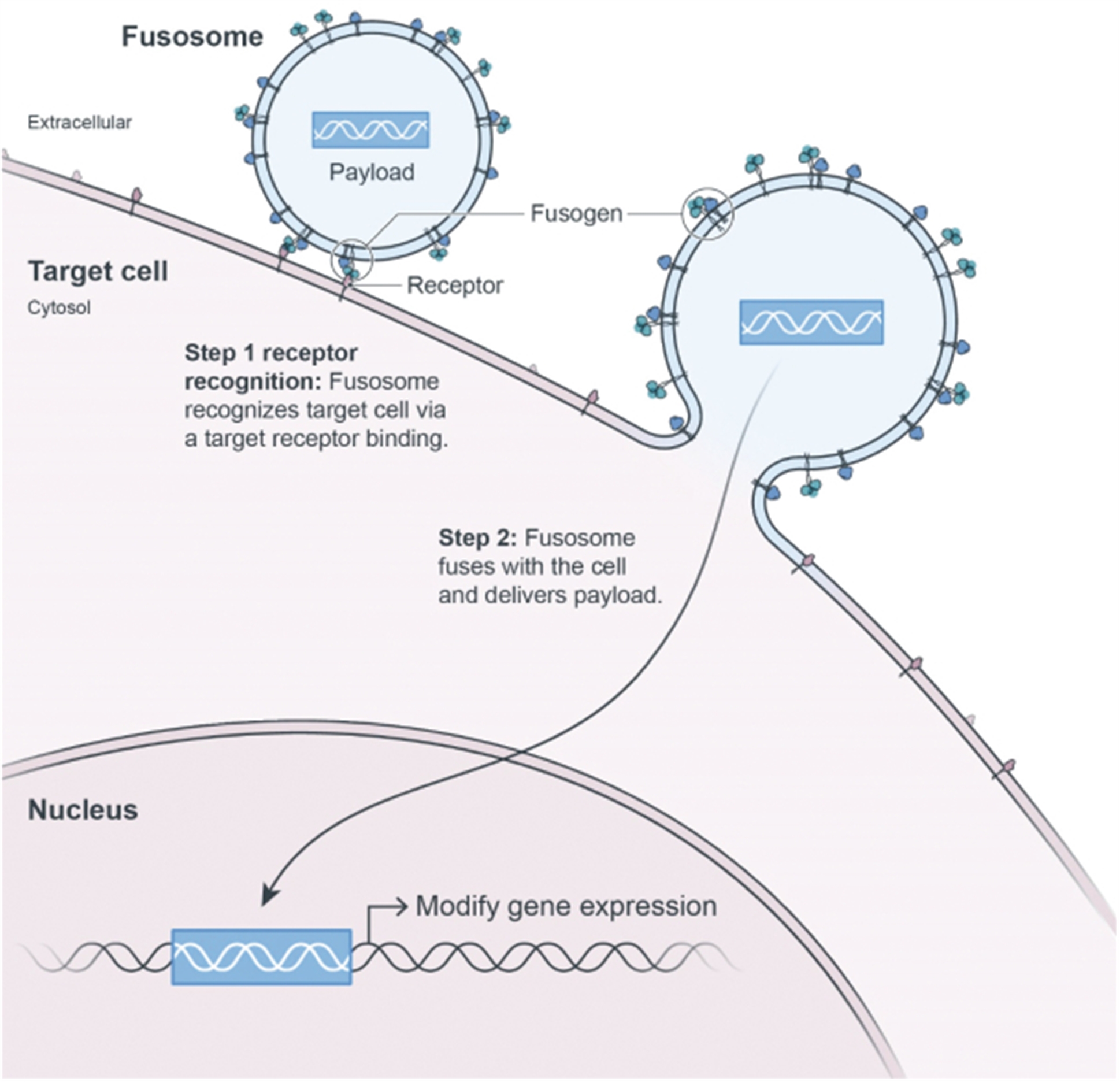
Fusogens are widely used by enveloped viruses to confer target specificity and to drive the process of introducing material in target cells. A well-known current example of a viral fusogen is the SARS-CoV-2 coronavirus that causes COVID-19. This virus uses its spike glycoprotein to target cells expressing the ACE2 receptor and to fuse with the cell membrane of host cells and release the viral genome into the cell. Many other biological processes utilizing fusogens for the delivery of complex, diverse, and large payloads to specific cell types have also been found. For example, the process of fertilization occurs as a result of a sperm fusing specifically with the egg and the transfer of the paternal genetic material to the oocyte. Similarly, the fusion of myoblasts with other myoblasts is essential for the formation, growth, and regeneration of skeletal muscle. The myoblast delivers an entire novel nucleus to the muscle cell, highlighting the utility of this system to deliver quite large and complex payloads. These and a myriad of other processes rely on this vast class of protein machines.
Applying fusogens to in vivo cell engineering
Building on both our team’s deep understanding of fusogen biology and extensive research in protein engineering, we are developing a technology designed to allow us to engineer the biological properties of these naturally occurring proteins. In doing so, we are developing a highly modular system that can specifically target numerous cell surface receptors and thereby deliver diverse therapeutic payloads to a variety of cell types.
40
Our current programs use fusogens derived from a virus from the paramyxoviridae family. The fusogen protein complex is comprised of two proteins: the receptor recognition G protein and membrane fusion F protein. The combination of a fusogen with a delivery vehicle such as a gene therapy vector or lipid vesicle is referred to as a fusosome. The diagram below depicts the mechanism of fusogen-mediated membrane fusion. This protein complex is found on the outer membrane of the fusosome (1). As the fusosome interacts with cells, only those with the target receptor will engage the G protein of the fusogen complex (2). The binding of the G protein to the receptor stimulates the F protein to initiate its membrane fusion activity. The F protein first partially unfolds to bind to the target membrane (3) and then refolds to bring the target and fusosome membranes in proximity (4), to ultimately promote membrane fusion (5), and subsequent payload delivery.
Mechanism of Fusogen-Mediated Membrane Fusion
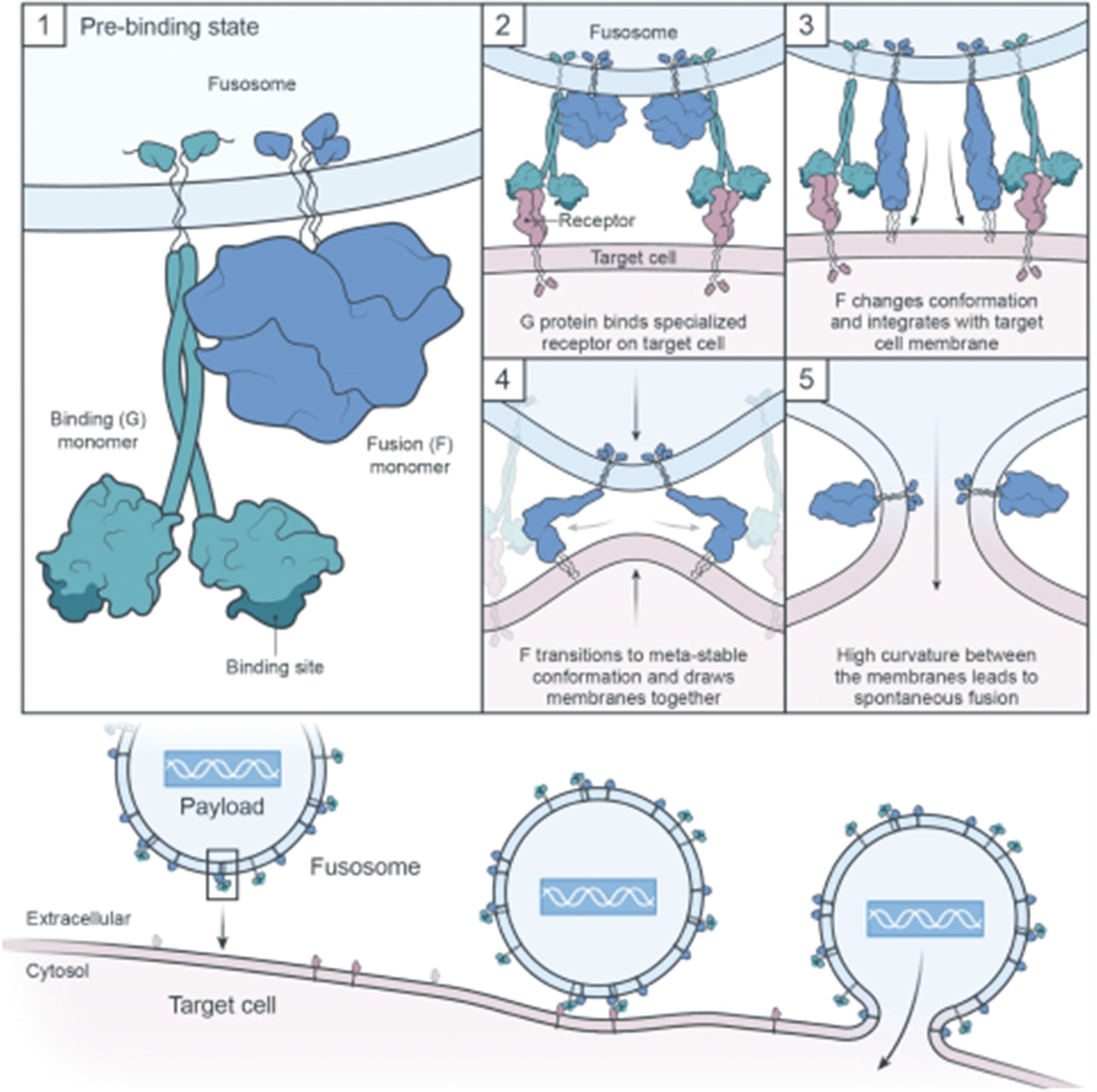
The G protein has the potential to be engineered for a high degree of cell selectivity. To accomplish this, we first engineer the G protein so that its natural binding domain is no longer functional. We then add a targeting scaffold to the G protein that re-directs the fusogen to a cell-specific receptor. The targeting scaffold can be any one of naturally occurring or synthetic single chain affinity binders, such as single chain variable fragment (scFvs), camelid single-domain antibodies (VHHs), or designed ankyrin repeat proteins (DARPins). Finally, we iteratively rebuild our fusogen using insights from protein engineering to improve titers, or potency. By serially swapping different targeting scaffolds we believe we can target multiple different cell surface receptors, giving us the ability to target many different cell types.
Re-targeting the specificity of the G-protein is a challenging protein engineering problem, since altering the protein structure directly impacts all aspects of biological function. However, once we have achieved the desired specificity and potency for a certain cell type, we have the ability to deliver a variety of payloads to that cell. This feature of the technology should allow us to create multiple therapies targeting a variety of diseases with each successful fusogen. As a result, we believe success with any initial therapy targeting a given cell type could meaningfully advance lead candidate selection for other indications and increases our confidence that we will be successful with subsequent therapies targeting that same cell type. For example, a successful hepatocyte-targeting fusogen applied to a fusosome for a given monogenic liver disease meaningfully accelerates lead candidate selection and increases our confidence that we will be successful with subsequent therapies targeting hepatocytes.
41
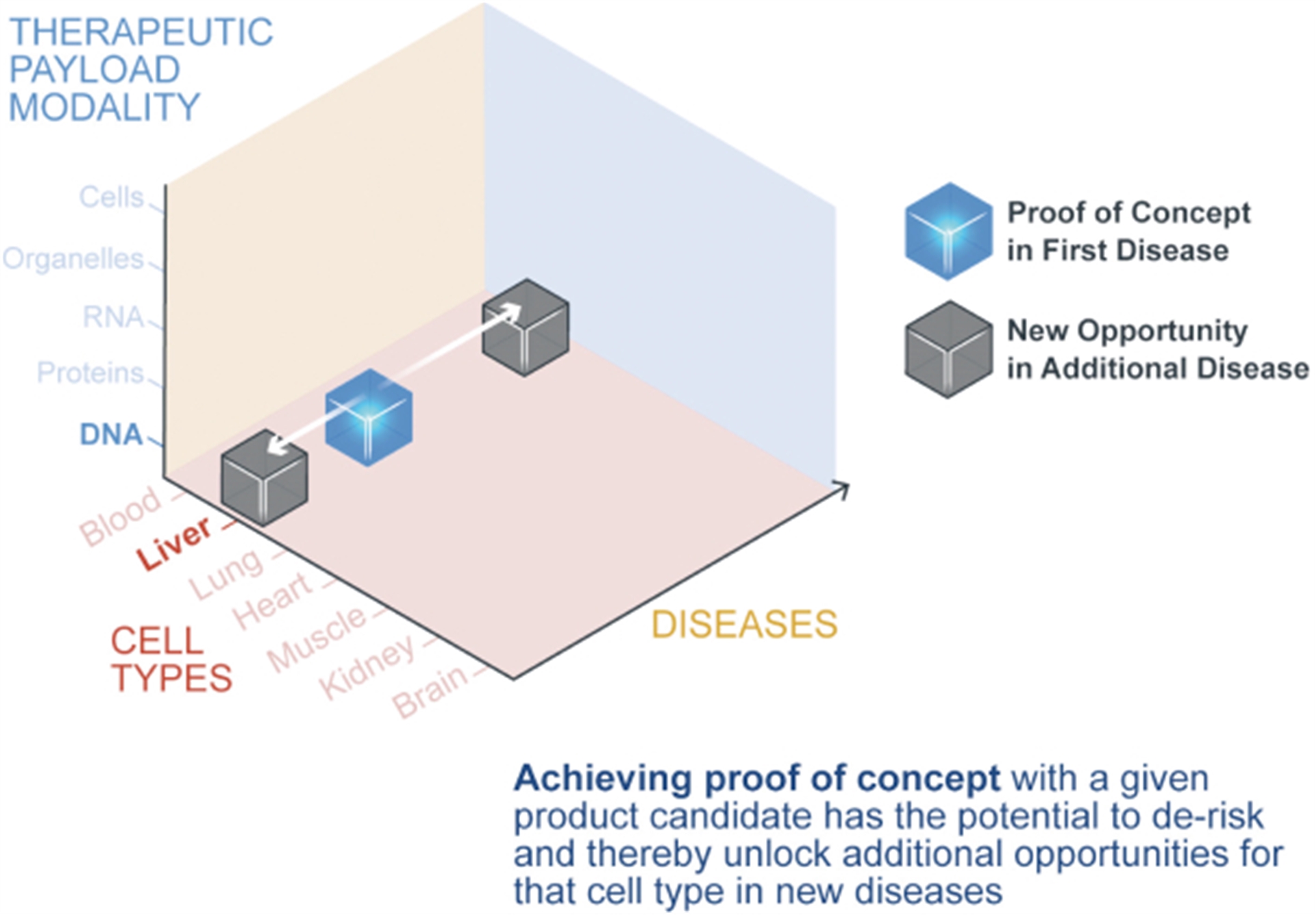
Addressing key in vivo cell engineering challenges
We believe that our in vivo cell engineering platform enables us to address key challenges associated with successful in vivo cell engineering – payload delivery, genome modification, and execution in manufacturing:
Payload delivery
High cell specificity for diverse cell types. We believe we can engineer fusogens with cell specificity to maximize on-target effects, while reducing or eliminating off-target risk. In our research, we have used fusogens to successfully target numerous cell surface receptors and cell types. As an example, in preclinical studies, we have demonstrated that our fusogens can specifically target CD8, CD4, or CD3 T cells (see the subsection titled “Our in vivo Cell Engineering Pipeline—T cell Fusosome Program”), potentially enabling delivery of a payload in vivo to transduce specific T cell populations and enabling targeted cell killing through the creation of CAR T cells.
42
Engineering Fusogens to Target a Variety of Cell Types
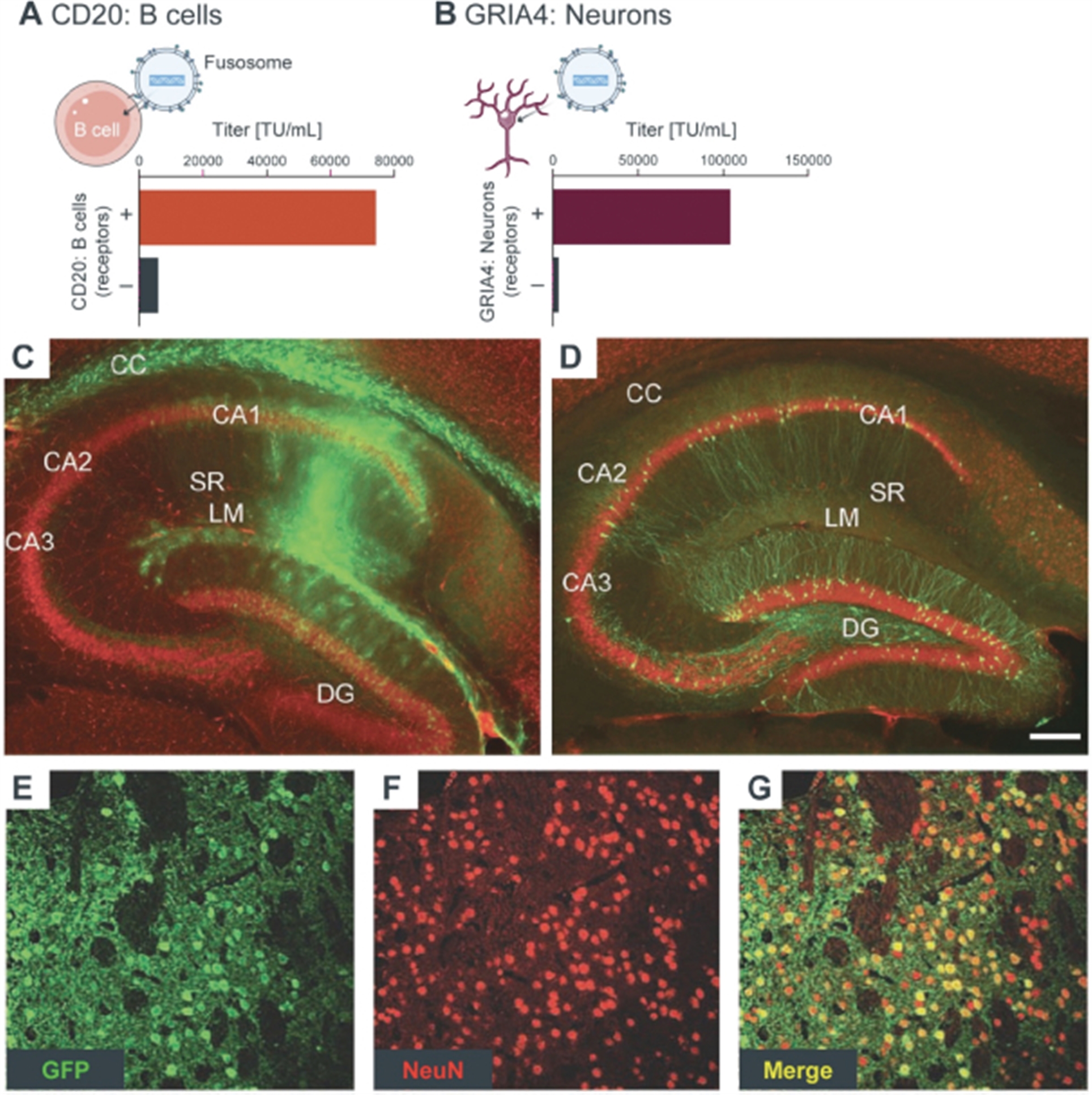
Figure A depicts the increased transduction efficiency (measured in titer) of a fusogen engineered for targeting CD20 on receptor enriched B cells as compared to B cells that were negative for the receptor. Similar transduction efficiency was also observed in an engineered fusogen targeting a neuronal surface protein, GRIA4, as depicted in Figure B. Neuronal-specific transduction of the Green Fluorescent Protein (GFP) payload in the murine hippocampal region was observed using a fusosome specific for GRIA4 when injected into the hippocampal space (as depicted by the green coloring in Figure D) compared to widespread transduction when using a VSV-G fusogen (Figure C). Confirmation of neuron-specific targeting of the fusogen can be observed by the colocalization of GFP positive cells (green, Figure E) with the presence of a neuron-specific protein (NeuN in red, Figure F) and considering the high degree of overlap (colocalization seen as yellow, Figure G). Figures C-G from Anliker et al, Nature Methods, 2010.
Broad volume of distribution. Our SanaX business unit is actively working on next generation approaches to broaden the volume of distribution, including exploring cells as fusosome delivery vehicles.
Immunogenicity. We have initially focused our efforts on selecting fusogens for which the general population does not have pre-existing immunity. We are also working with a number of fusogens that exist naturally in humans, as neither these native fusogens nor re-targeted versions are likely to induce an immune response, making re-dosing more readily attainable.
43
Genome modification
High degree of payload flexibility. We have successfully delivered a variety of payloads including DNA, RNA, and proteins, using viral delivery methods and have used cells engineered to express specific fusogens to deliver organelles to a broad range of target cells. We believe this provides us the opportunity to potentially intervene in a wide range of human diseases.
Diverse Payload Delivery via Fusosomes
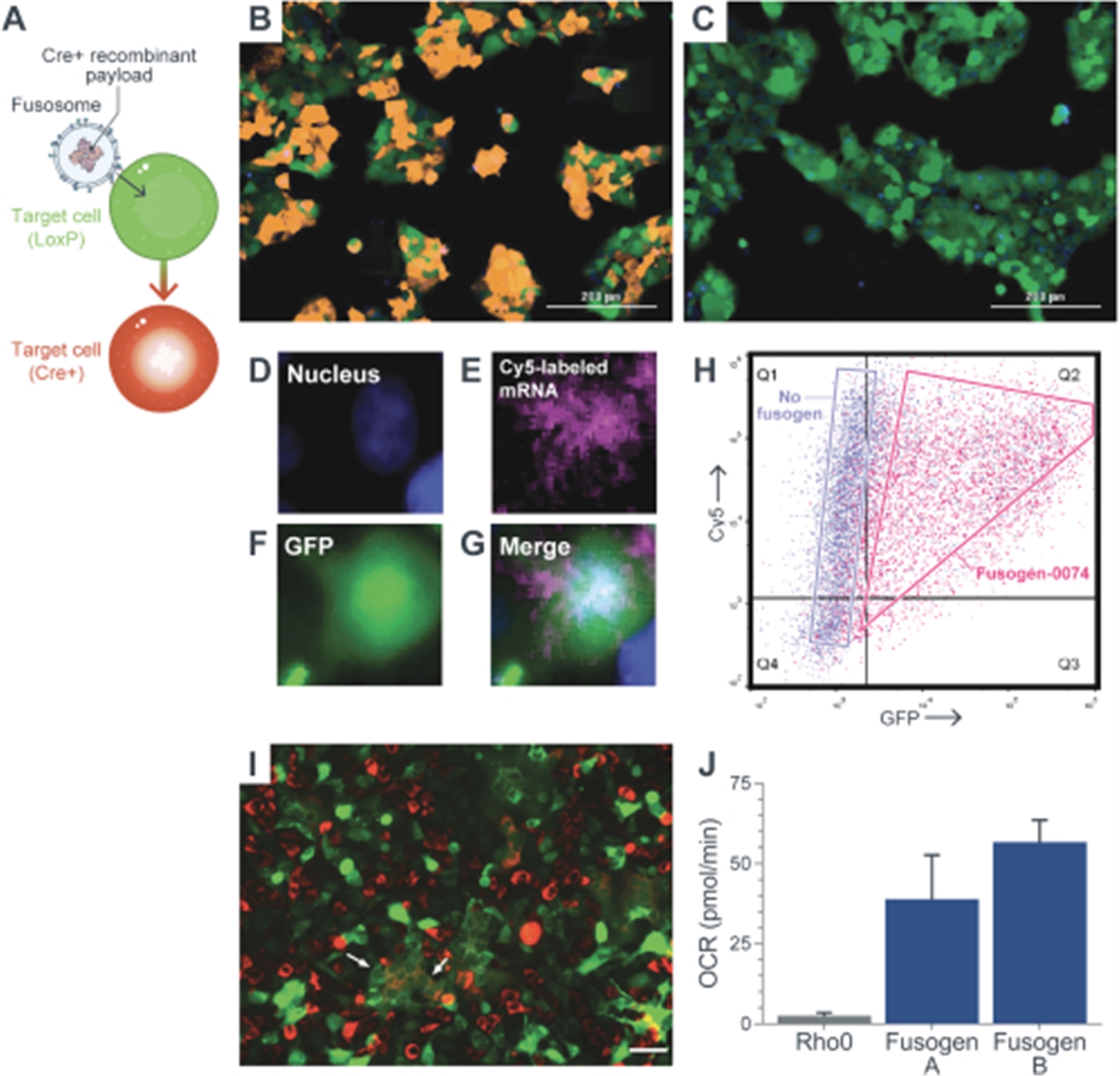
Cre protein loaded cell-based fusosomes delivered recombinase activity to cells that activated the expression of a red fluorescent protein in cells already expressing green fluorescent protein, seen as orange cells (Figures A, B). In contrast, fusosomes in which the fusogen is not included, but only contain Cre protein, showed no recombinase activity, or no orange cells (Figure C). Fusosomes loaded with fluorescently labeled RNA showed cellular localization and green fluorescence consistent with cytoplasmic delivery and translation of delivered RNA (Figures D-G). Flow cytometric analysis showed cellular uptake of fluorescent RNA (Cy5, Y axis) and GFP expression from the RNA (GFP, X axis) (Figure H). Importantly, the inclusion of a fusogen in the fusosome dramatically increased GFP expression due to the translation of the RNA. Cell-based fusosomes delivered red fluorescent mitochondria with respiration activity to cells with respiration-negative green mitochondria, (Rho0 cells) shown in Figure I. An increased oxygen consumption rate (OCR), due to respiration, was seen in Rho0 cells after Fusosome-mediated delivery of active mitochondria using two distinct fusogens (Figure J).
Expanded payload capacity. Our current fusosome has approximately twice the genetic capacity of the commonly used AAV vectors. This greater payload size increases the potential of addressing defects in larger genes or conditions where delivery of multiple genes may be required. Our research efforts include other fusosomes with even larger payload capacities. For example, utilizing a cell as the delivery vehicle can confer an almost limitless capacity.
Durability limitations. We can engineer our fusosomes to integrate into the target cell genome or to deliver non-integrating payloads. Integrated payloads allow the genetic information transmitted by the vector to be propagated durably with the genetic material of the target cell when it undergoes cell division. Thus, conditions that require this type of genetic propagation, such as genetic diseases in essential genes functioning in growing tissues or in T cells expanding after recognizing a target antigen, can be better addressed by this approach. Our preclinical studies have also demonstrated the ability to deliver gene-editing machinery, such as CRISPR, with this system. In this case, the entire payload does not integrate, but instead, it transiently delivers the machinery to
44
permanently modify the DNA in the target cell. Thus, we are able to make targeted, specific, and durable repairs to the genome of the target cell.
Execution in Manufacturing
Manufacturing of cell and gene therapies remains complex due to incumbent challenges in areas such as product consistency, process robustness, and scalability. Our fusosome approach has significant advantages over current solutions. Targeted delivery of complex payloads in vivo has the potential to create autologous, gene-modified cells without the complexities of ex vivo manufacturing. We believe that these therapies have the potential to have greater product consistency, improved scale, and lower costs than current autologous solutions. Currently, there are a number of therapies either approved or in development for ex vivo modification of autologous T cells and autologous HSCs. Additionally, vectors that deliver payload to random or off target cells not only create the risk for toxicities, but they necessitate meaningfully larger doses in order to ensure adequate delivery to the targeted cells. Our targeted delivery offers the potential for meaningfully lower doses, which could decrease scale needs in manufacturing.
Further, we are investing across a number of areas to improve manufacturing scale, costs, consistency, and product quality in the near-term and long-term, including by establishing and maintaining our relationships with our CDMO partners and investing in establishing and operating our own GMP manufacturing facility. Manufacturing novel fusosome compositions is complex. Since our inception, we have invested in scientific and process engineering expertise to improve manufacturing of our therapies. Examples include novel producer cell lines, novel processes and analytical technology, as well as incorporating suspension bioreactors into our process early in the research phase. By building out these capabilities early, we hope to improve the probability of technical success for our programs and have a thoughtful approach to deliver consistent supply while managing cost of goods with the goal of improving patient access.
Our in vivo Cell Engineering Pipeline
T Cell Fusosome Program (SG242, SG295, SG233, SG221, SG239)
Our most advanced CAR T cell fusosome product candidates (SG242, SG295) target CD19+ cancer cells, including NHL, CLL, and ALL. We intend to develop these product candidates with the goal of submitting an IND as early as 2022. In parallel with the CD19 CAR product candidates we are developing other CAR T cell therapies, including BCMA product candidates for the treatment of multiple myeloma (SG221, SG239), CD22 product candidates for the treatment of NHL, CLL, and ALL (SG233), as well as other targets on a spectrum of cancers.
Background on B Cell Malignancies
B cell malignancies represent a spectrum of cancers including non-Hodgkin lymphoma (NHL), chronic lymphocytic leukemia (CLL), acute lymphoblastic leukemia (ALL) and multiple myeloma (MM) and result in over 100,000 deaths per year in the United States and Europe.
NHL is the most common cancer of the lymphatic system. NHL is not a single disease, but rather a group of several closely related cancers. Over 77,000 cases of NHL are diagnosed annually in the United States and the most common subtype of NHL overall is diffuse large B cell lymphoma (DLBCL). DLBCL, if left untreated, may have survival measured in weeks or months.
CLL is the most common type of leukemia, and it occurs most frequently in older individuals, with diagnoses in people under 30 years of age occurring only rarely. Each year, approximately 20,000 patients are diagnosed with CLL in the United States. Approximately 20–25% of CLL patients initially present with poor risk disease. Median progression-free survival in these high-risk groups is often less than 12 to 18 months after frontline therapy, and less than 12 months in R/R disease.
ALL is an uncontrolled proliferation of lymphoblasts, which are immature white blood cells. The lymphoblasts, which are produced in the bone marrow, cause damage and death by inhibiting the production of normal cells. Approximately 6,000 patients are diagnosed with ALL in the United States each year, the vast majority of the approximately 1,500 deaths per year occur in adults. Approximately 80% of cases of ALL in the United States and Europe are B cell ALL, which almost always express the CD19 protein. The five-year overall survival in adults over the age of 60 with ALL is approximately 20%, and in patients with R/R ALL after two or more lines of therapy, the median disease-free survival is less than six months. B cell ALL is the most common cancer in children. Although children with ALL fare better than adults, children with R/R disease have poor outcomes. Because of the frequency, ALL remains a leading cause of death due to cancer in children.
Multiple myeloma is a cancer of the plasma cells, which typically express a protein called B Cell Maturational Antigen (BCMA). Plasma cells are B cells that have matured to specialize in the production of antibodies. Multiple myeloma is a condition in which these plasma cells become malignant, with a single clone growing at an uncontrolled pace. These myeloma cells secrete large
45
quantities of the same antibody, and patient symptoms can develop from the myeloma cells crowding out other plasma and bone marrow cells, leading to increased risk of infection, risk of bone destruction, and kidney disease. Multiple myeloma is the second most common hematologic malignancy making up approximately 2% of all cancers, accounting for over 32,000 new cases per year with 12,800 deaths estimated to occur in 2020 in the United States.
Current Treatment Landscape and Unmet Need
First line therapy for NHL typically consists of multi-agent cytotoxic drugs in combination with the monoclonal antibody Rituxan. In younger patients with NHL who have good organ function, high dose chemotherapy followed by stem cell transplantation is often used. Patients often relapse, however, and over the last three years, several therapeutics have been approved in the United States for the treatment of patients with R/R NHL who have received prior therapies. These approved therapies include CD19 CAR T therapies tisagenlecleucel, axicabtagene ciloleucel and lisocabtagene maraleucel, CD19 antibody drug conjugate therapy polatuzumab vedotin, and CD19 antibody tafasitamab. Recently, pivotal trials with approved CD19 CAR T cells have been shown to be superior to standard of care approaches in patients in second line R/R NHL raising the possibility of broader impact for CD19 CAR T cells for patients with NHL.
Newly diagnosed CLL patients are often treated with targeted therapies such as BTK inhibitors, PIK3 inhibitors, BCL-2 inhibitors, or monoclonal antibodies targeting CD20, or CD52 in combination with chemotherapy. However, most patients treated with these regimens become refractory. Numerous drug candidates are in clinical development for the refractory patients, including next-generation kinase inhibitors and both autologous and allogeneic CAR T therapies targeting CD20 and CD19.
Cure rates for ALL patients have continued to increase over the last four decades, with pediatric ALL cure rates reaching greater than 80% in developed countries. This progress has been enabled by advances in combination chemotherapy, monitoring of minimal residual disease, expanded use of kinase inhibitors for Philadelphia chromosome–positive ALL, and the recent approval of Kymriah for R/R pediatric ALL. Adult patients fare much worse, however, with 5-year overall survival rates of approximately 20%, and there are still significant challenges managing R/R disease across all age groups. Multiple therapeutic candidates are in development for these R/R patients, including proteasome inhibitors, antimetabolites, JAK inhibitors, monoclonal antibodies, as well as autologous and allogeneic CAR T candidates.
First-line therapy for MM is induction and high-dose chemotherapy followed by a potential stem cell transplant. There are no curative treatment options for MM patients and the standard of care for R/R MM includes immunomodulary agents, proteasome inhibitors, monoclonal antibodies, cytotoxic agents, and hematopoietic stem cell transplant. Despite the recent advancement in available therapies for disease management, the 5-year overall survival rate remains approximately 50%. To this end, several groups are investigating autologous and allogeneic CAR T cell therapies for R/R MM. BCMA is among the most promising antigens used to target MM, with two approved BCMA CAR T therapies idecabtagene vicleucel, and ciltacabtagene autoleucel, as well as multiple late-stage clinical trials ongoing. Novel treatments with other mechanisms of action are also under development, including bispecific T cell engagers, next-gen antibodies, and antibody drug conjugates.
As highlighted above, recent therapeutic advances across R/R B cell malignancies have led to a variety of treatment options and better patient outcomes. In particular, autologous surface protein directed CAR T therapies have been highly effective in certain subsets of patients with R/R disease. However, not all patients have access to novel therapies, and even with them, many patients will ultimately relapse and succumb to their cancer, resulting in 100,000 deaths per year in the United States and Europe across these indications.
There are two outstanding challenges that have limited utilization of these CAR T therapies and their impact on broader groups of patients.
Relapse. The emerging post-approval data with tisagenlecleucel and axicabtagene ciloleucel have indicated that there are two broad categories of relapse. One involves loss of CD19 on malignant cells resulting in tumor escape. This finding was initially established for ALL and is the cause of relapse after CAR T cells for roughly half of patients. More recent data indicate that low antigen expression contributes to the lack of response in a meaningful number of patients with NHL. CD19 CAR T treatments have recently been tested in pivotal trials in earlier lines of therapy for NHL, which raises the possibility of more patients being treated with CD19 CAR T therapy who subsequently relapse due to CD19 loss. Therefore, the development of CAR Ts targeting an alternate antigen beyond CD19 may be beneficial to address this growing unmet need. Data from several studies have shown that CD22 CAR T treatment has led to complete responses in NHL and ALL patients that have failed to reach a complete response or relapsed after treatment with a CD19 CAR T cell product. Additionally, dual targeting of CD19 and CD22 as the initial form of treatment may prevent this form of relapse, offering patients the potential for both a higher rate and longer duration of complete response. The second pattern of relapse relates to suboptimal CAR T cell functionality (poor expansion, poor persistence, T cell exhaustion) resulting in relapse of cancer that retains the targeted antigen. Unfortunately, re-infusion of the same CAR T cell product has had limited benefit
46
in these patients although treatment with a different CAR T cell has demonstrated some promise in the context of ongoing clinical trials.
Manufacturing. The manufacturing process for a patient-specific product is complex, leading to limited access due to both infrastructure and cost considerations. As such, approved CAR T cell therapies have not been available to all patients in need of these highly effective therapies. Even for patients who are fortunate enough to have access, inevitable delays (often a month or more) in manufacturing may prevent use of therapy in patients with rapidly progressing malignancies. There are groups that are seeking to overcome access limitations by using healthy donor-derived, or allogeneic, CAR T cells instead of patient T cells. This approach yields off-the-shelf therapeutics that can be manufactured consistently, but questions remain around efficacy and durability, largely due to the inability to effectively control the host versus graft response with concern for eventual rejection of these products. As will be discussed in the subsection titled “—Our ex vivo Cell Engineering Pipeline,” our ex vivo allogeneic T cell program also seeks to address this host versus graft response.
T Cell Fusosome Program
Our T cell fusosome approach provides us with an opportunity to develop potential product candidates to expand access to CAR T cell therapy to many more patients in need. In addition, we believe the ability to deliver a payload encoding a CAR to a T cell inside the body has the potential to improve effectiveness over ex vivo manufactured CAR T cell products. Experience thus far has demonstrated that both CD8+ and CD4+ T cells contribute to the CAR T cell response. Thus, the fusosome programs we are developing will deliver the CAR gene using fusogens that directly and specifically target the CD8 co-receptor or the CD4 co-receptor on T cells following a single intravenous injection. These approaches could result in the generation of therapeutically active CAR T cells without the complexities and delays associated with the process of T cell collection and ex vivo manufacturing. Furthermore, the ex vivo expansion in the presence of high cytokine concentrations, while necessary for the manufacture of approved CAR T cell products, also contributes to marked changes in T cell quality that may not be therapeutically beneficial. We believe the generation of a CAR T cell within the natural physiological environment has the potential to improve the quality of the CAR T cell generated, potentially improving both efficacy and the side effect profile. Finally, the effectiveness of ex vivo manufactured CAR T cells is dependent on the administration of a lymphodepleting preparative regimen prior to infusion to facilitate expansion of the CAR T cell product, which can have meaningful adverse safety implications. We do not expect to need a lymphodepleting regimen prior to in vivo delivery of the CAR gene, as our goal is to expose our fusosomes to as many T cells in the body as possible.
Preclinical Data
Our preclinical data have demonstrated that fusosomes can deliver a genetic payload specifically and efficiently to human T cells in culture and in immunodeficient mice with intraperitoneally injected human peripheral blood mononuclear cells (PBMC) and fused with a single dose of a fusosome. The T cells can be categorized into functional subsets based on the expression pattern of cell surface molecules. CD3 is a protein expressed on all T cells, CD4 is expressed on the Helper T cells that primarily activate T and B cells to carry out their function, and CD8 is found on cytotoxic T cells that primarily kill cancerous or virally infected cells. We generated fusogens against these three cell-surface molecules and have demonstrated that we can deliver a marker gene to cells bearing these cell surface proteins in vitro.
47
Fusogens Demonstrate the Ability to Target Multiple T Cell Subtypes
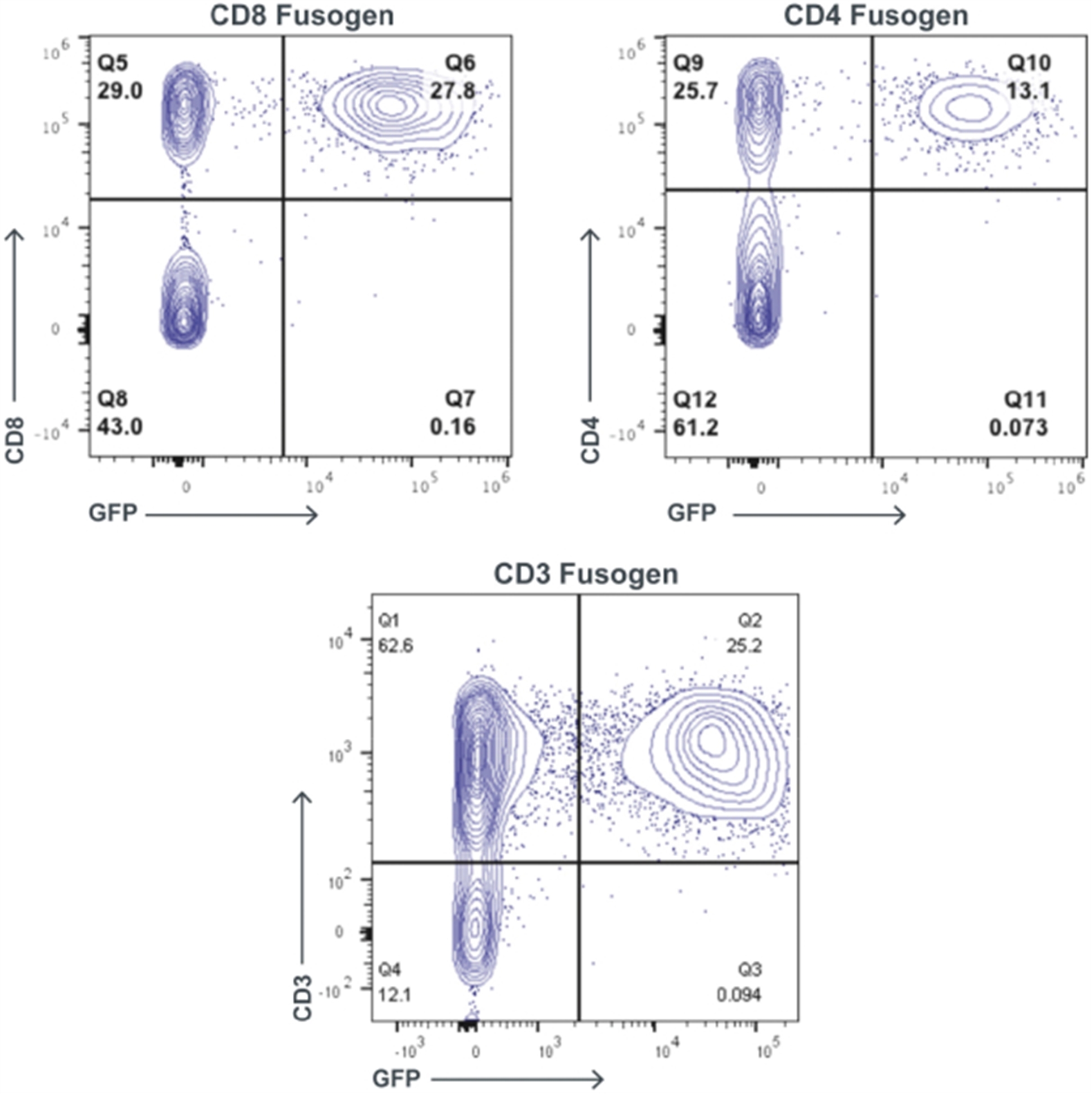
Fusosomes containing a gene that encodes a fluorescent marker protein called GFP (used to identify cells have been genetically modified by the fusogen) can efficiently and specifically deliver GFP to T cells in culture (CD8, CD4, and CD3). Expression of GFP is restricted to the population of T cells that express the specific T cell receptor targeted by the fusogen (CD8, CD4, or CD3).
48
We have further established that fusosome delivery of a CD19 CAR gene to CD4 or CD8 T cells results in killing of human B cells and CD19+ leukemia cells in culture:
Delivery of CD19 CAR to CD4 T Cells Leads to in vitro Killing of B Cells and CD19+ Leukemia Cells
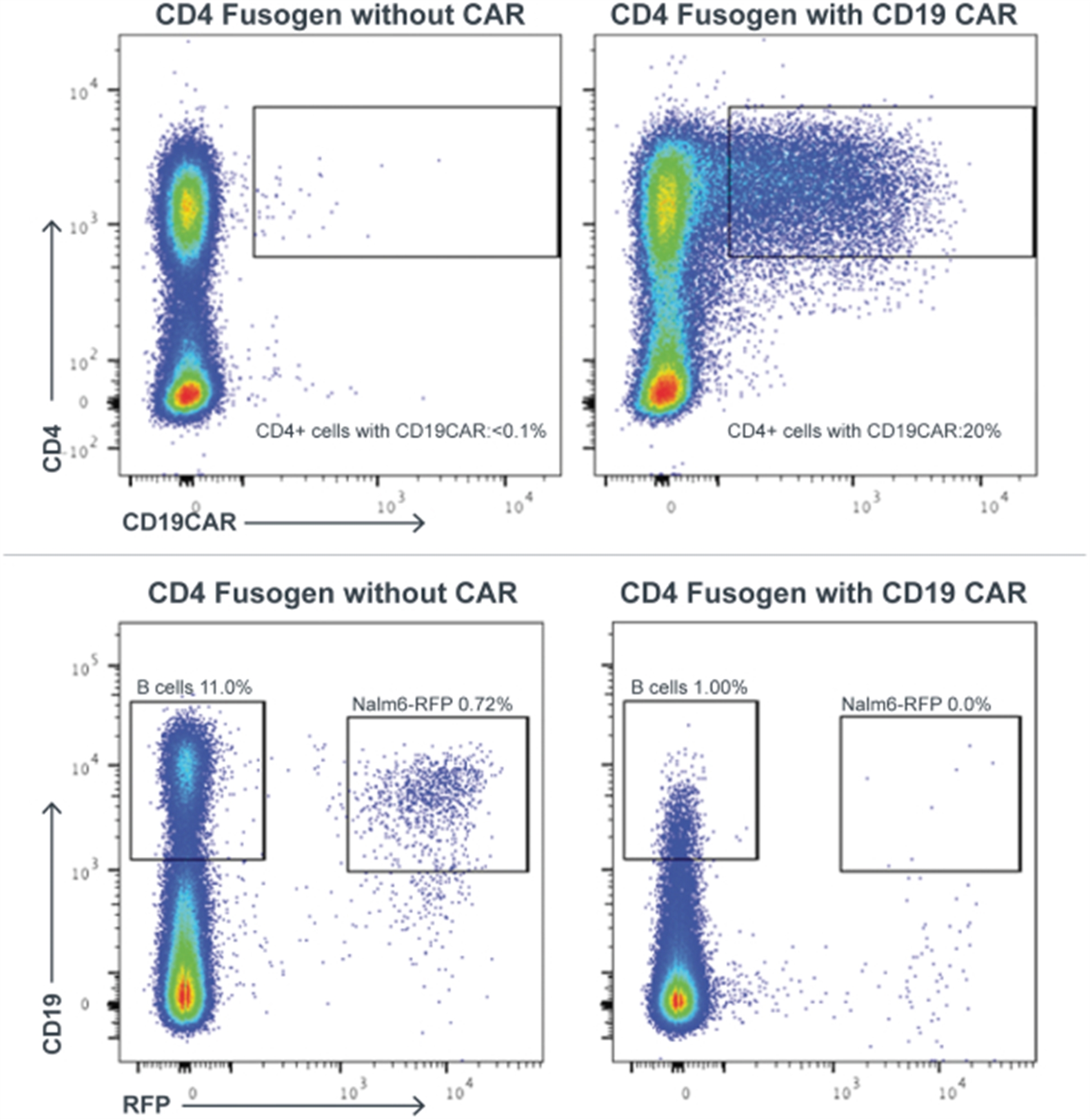
Demonstrates that the fusosome-generated CD4 CAR is functional and eradicates both nonmalignant B cells (CD19+/RFP-) as well as CD19+ leukemia cells expressing NALM6-RFP.
We have also validated, in vivo, the tumor-killing activity of CD8 T cells to which CD19 CAR has been delivered via a fusosome.
49
Delivery of CD19 CAR to CD8 Cells Leads to in vivo Killing of Leukemia Cells in a Human Xenograft Mouse Model
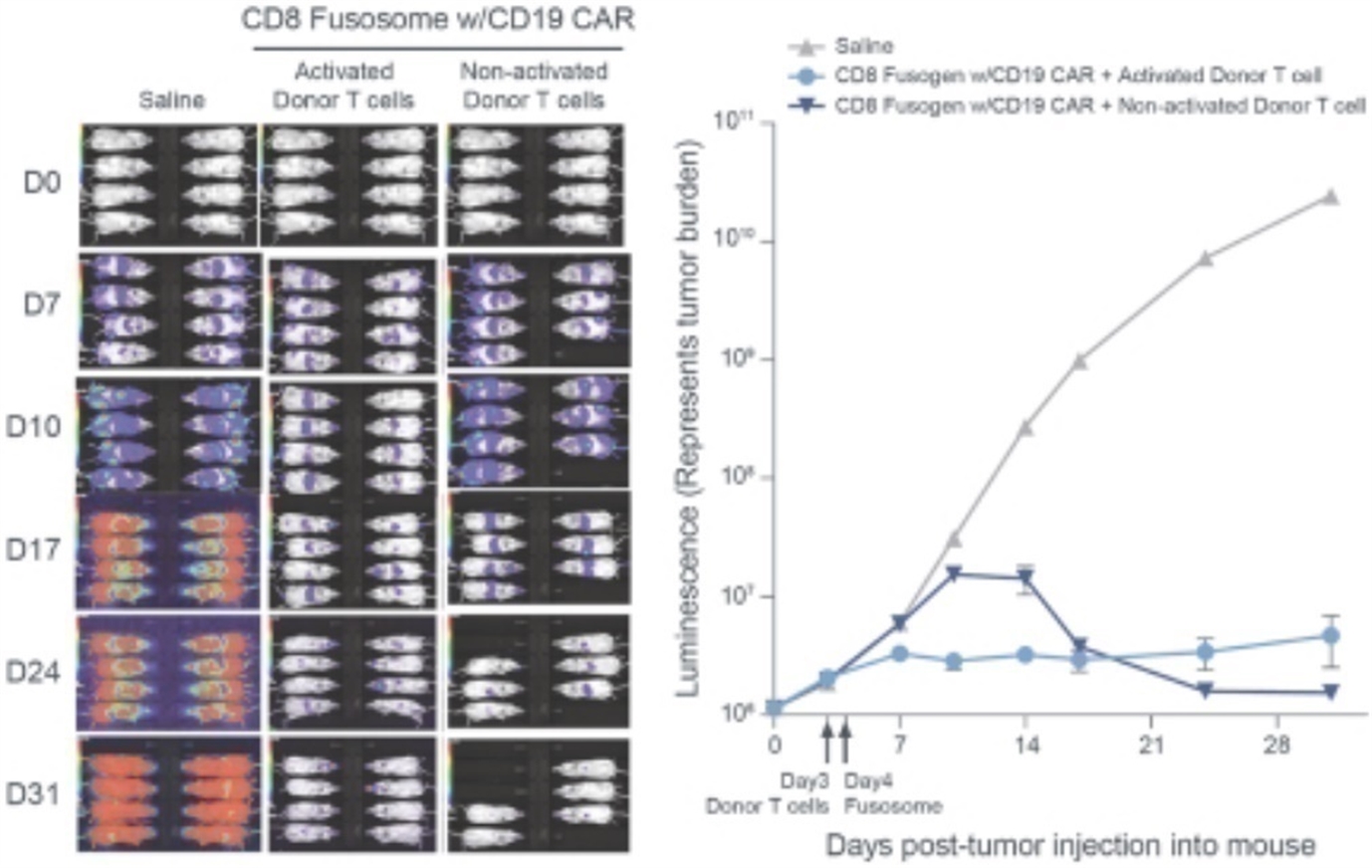
Left panel: demonstrates activity of CD8 fusosome delivering CD19 CAR to human T cells in a murine leukemia xenograft model (Nalm-6). Note that when compared to untreated controls, fusosome delivery results in eradication of leukemia cells. Activated T cells were cultured with CD3/CD28 beads for 3 days prior to injection. CD8 fusosome delivering the CD19 CAR is effective regardless of activation status of T cells at time of injection. Right panel: represents quantification of luminescence (representing leukemic burden) from mice shown in left panel. Both cohorts of fusosome treated mice had significantly reduced tumor burden when compared to control as early as D10 (p≤.0001; One-way ANOVA Bonnferroni) Experimental note: Tumors injected on Day Zero, Donor T cells injected on Day Three and Fusosome injected on Day Four.
Using a human xenograft mouse model for leukemia (Nalm-6), we observed both prolonged survival and clearance of the leukemic cells. During the manufacture of autologous CAR Ts, cytokine signaling has to be activated in order to successfully produce functional CAR T cells. In our mouse experiments the CD8 fusosome was able to generate CD19 CAR cells just as effectively with activated as non-activated donor T cells.
Several of our human T cell fusogens cross-react on non-human primate (NHP) T cells including our lead candidate CD8 fusogen. We have used the fusogen to deliver a CD20 CAR into six NHPs (the CD20 CAR was chosen as the CD19 CAR to be used for our clinical programs does not cross-react with NHP B cells). As shown below, a single intravenous administration of our CD8 fusogen containing a CD20 CAR was associated with B cell depletion, including in the blood and in lymph nodes, in four out of six NHPs, occurring between day seven and fourteen. This result is consistent with reported observations that peak expansion of ex vivo manufactured CAR T cells typically occurs during the second week. Serum cytokines were transiently elevated at day seven in all fusosome-treated animals. CD20 CAR transgene (by vector copy number) and CD20 CAR mRNA could be detected in peripheral blood between days three to ten, and in the spleen at study termination. Importantly, there was no infusion-related toxicity or evidence for CAR-associated toxicity (cytokine release syndrome or neurotoxicity) other than the intended B cell depletion. The ability to deliver fusogen without toxicity and with evidence for activity in NHP are critical milestones for the program. In addition, the NHPs received no T cell activating agent or lymphodepletion. The latter potentially supports a path for the fusogen platform to enable delivery of CAR therapy without the lymphodepletion regimens used by existing ex vivo approaches, which have toxic side-effects. This and future NHP experiments will also provide important information on dosing parameters, durability of the effect, and provide pharmacokinetic, pharmacodynamic, and toxicology data.
50
Delivery of CD20 CAR to CD8 Cells Causes B Cell Depletion in NHPs
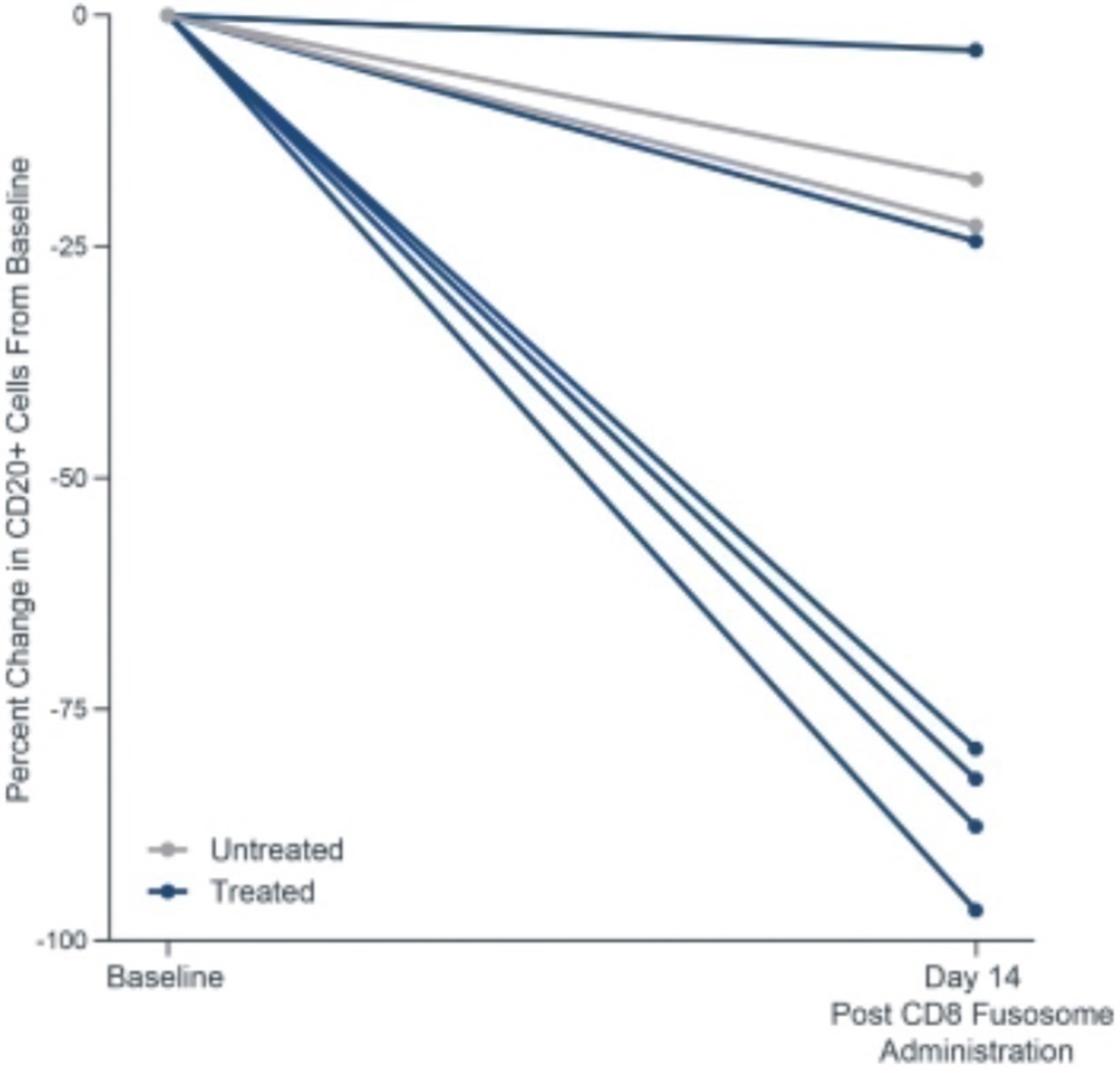
Graph demonstrates activity of CD8 fusosome delivering CD20 CAR in NHPs. CD8 fusosome was delivered as a single intravenous infusion. B cell counts were quantified in peripheral blood using flow cytometry for CD20+ cells. Given expected NHP-to-NHP variability in baseline counts, data is represented as single lines per NHP and is shown as deviation from baseline level for each NHP at Day Fourteen (when maximal expansion of CAR T cells are expected). Significant B cell depletion is observed in four out of six NHPs. Note that no T cell activation was provided prior to fusosome delivery.
Development Plan and Key Next Steps
We are currently conducting experiments to validate the ability of a systemically administered fusosome to transduce T cells in an NHP and for these CAR T cells to deplete B cells. These NHP studies are also expected to inform preclinical pharmacology and toxicology.
As a next step, we intend to conduct our in-life GLP toxicology studies. While these studies are ongoing, we intend to scale our GMP manufacturing and finalize our initial development plan. We intend to submit an IND for SG295 to support clinical trials in patients with NHL in 2022. We continue to advance additional programs and plan to submit INDs over the next several years: SG242 in NHL, SG233 which delivers a CAR gene targeting CD22 and could be combined with targeting CD19, and our BCMA programs in MM.
Hepatocyte Fusosome Program
Numerous genetic metabolic diseases arise from gene defects that manifest in the liver and, in particular, in the hepatocyte. Additionally, hepatocytes can serve as protein manufacturing sites to deliver proteins to other cells in the body. Multiple modalities exist that enable delivery of genetic material to liver cells, including AAV and LNPs. However, these approaches have limitations, including non-integrating payloads, payload size, lack of cell specificity, and, in the case of AAV, immunogenicity. Our fusogen technology, which we expect will be able to deliver a payload specifically to hepatocytes in the liver, has the potential to address these limitations. Success with this hepatocyte-targeting technology may allow us to generate therapies for a number of genetic disorders. We are developing our lead product candidate, SG328, for ornithine transcarbamylase (OTC) deficiency, and we expect to submit an IND in the next several years.
Hepatocyte Targeting Capability
Targeting the hepatocyte with a fusogen can enable specific delivery of either integrating or non-integrating payloads. It can also be used to deliver the machinery of gene editing and gene modification tools to these cells, both with or without the inclusion of DNA to replace a mutated gene or gene fragment. Since we anticipate that hepatocytes transduced with fusosomes will harbor the novel genetic construct in their genome, all progeny of that cell will also have the genetic construct. Thus, the natural turnover and organ growth will not dilute the genetic construct, providing the potential for long-term expression and efficacy even when the
51
fusosome is delivered during infancy, childhood, or when it is delivered to treat a disease where the disorder can cause rapid hepatocyte turnover.
We believe that success with an initial hepatocyte-targeted fusosome will meaningfully accelerate our future hepatocyte programs. Once a hepatocyte-targeting fusosome is established, our subsequent programs will require only substituting the relevant payload to correct for the defective gene in question, opening up the possibility to address multiple inherited liver diseases. Our initial focus is on monogenic diseases with clear biology linking the missing activity of a gene in hepatocytes to a disease outcome. According to the National Institute of Health, over 30 genetic disorders of the liver exist, impacting over 10,000 births annually around the world. Many of these disorders lead to death in the first few years or cause long term disabilities. Proof of concept within this set of initial diseases will enable expansion to other diseases, such as hemophilia, where we may be able to address an unmet need by providing a durable in vivo therapy in the hepatocyte.
Preclinical Data
Our ability to use our hepatocyte-targeting fusosomes in relevant animal models is limited by a lack of cross-species transduction. To address this, we first developed murine disease models and introduced the therapeutic payload utilizing a conventional lentivirus pseudotyped with VSV-G. The VSV-G fusogen targets the LDL receptor which is highly expressed in hepatocytes providing a potent in vivo delivery vehicle for hepatocytes. These models established proof of concept to treat these diseases through genomic integration of the corrected gene as well as transduction efficiency in the range that would be needed for efficacy. However, the LDL receptor is found on a significant number of other cell types resulting in extensive off-target transduction. We do not intend to move forward with this non-specific lentivirus construct, as its lack of specificity creates potential challenges in humans. However, it does provide a preclinical model system for us to understand the percentage of hepatocytes, as well as the expression level of the novel genetic material that are required for the intended therapeutic effect.
In parallel, we have developed and improved hepatocyte-specific fusosomes for high on target transduction efficiency (as measured by titer), with the goal of achieving potency comparable to or better than what we see with conventional lentivirus. Engineering of hepatocyte specificity is generated through the choice of target receptor selectively expressed in human hepatocytes. Through an iterative process focusing on multiple hepatocyte-selective cell surface protein targets, diverse binders, and protein engineering, we have developed constructs that have met our potency goals. We have tested these constructs in vivo in mouse models and shown that they can transduce human hepatocytes at levels comparable to conventional lentivirus with significantly lower frequency of off-target transduction. Furthermore, the transduction occurs in a dose-dependent manner.
52
Improved Targeted Fusogens Exhibit High in vitro Transduction of Primary Human Hepatocytes (PHHs)
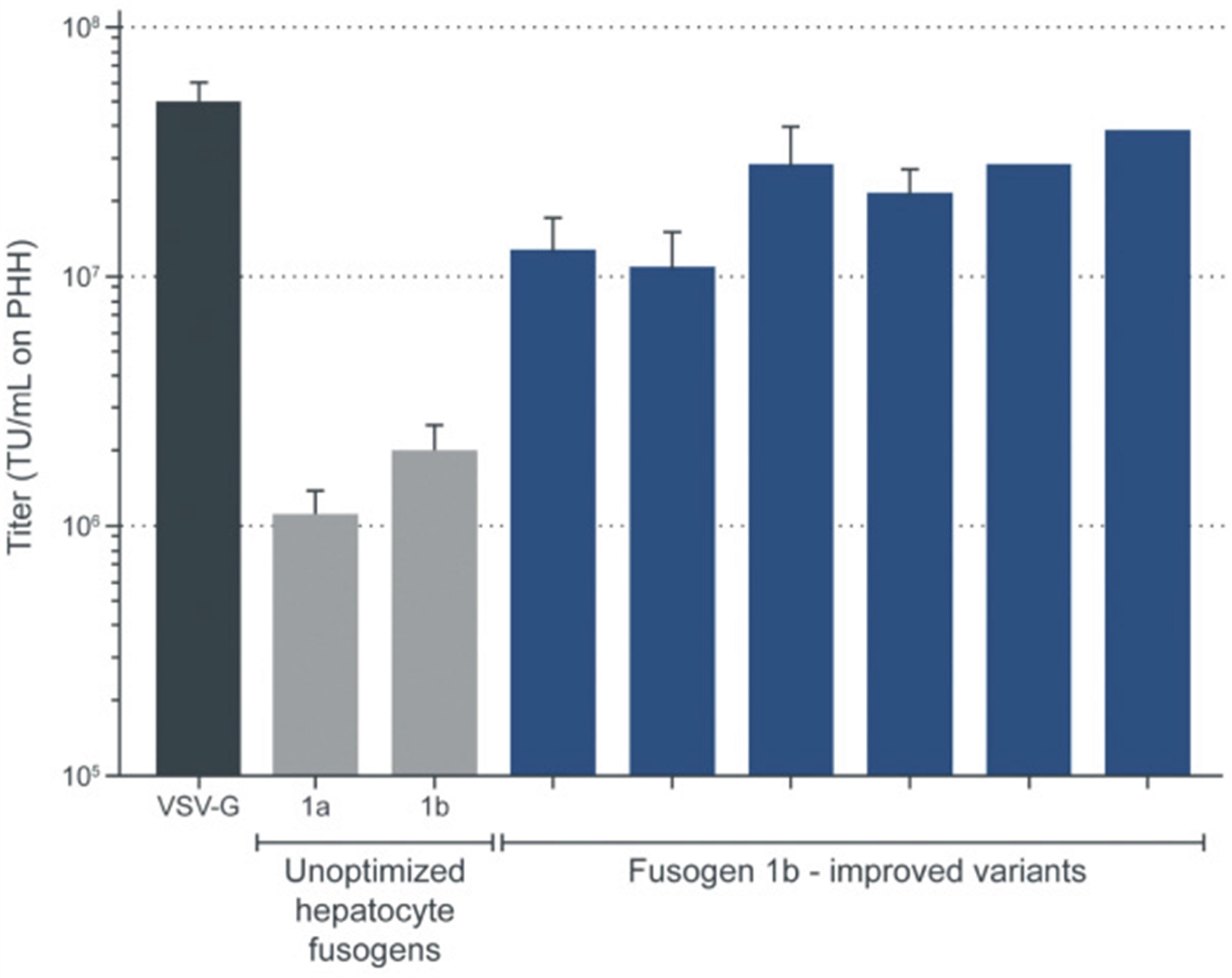
Improved hepatocyte-targeted fusosomes show levels of in vitro transduction similar to conventional lentivirus. Protein engineering of Fusogen 1b resulted in new sets of fusogens with significantly increased titer on PHHs. The most potent of these fusosomes approach the hepatocyte titer of conventional lentivirus.
53
Improved Targeted Fusogens Can Transduce Hepatocytes in vivo in a Humanized Liver Mouse (FRG) at Comparable Levels to Conventional Lentivirus and in a Dose-Dependent Fashion
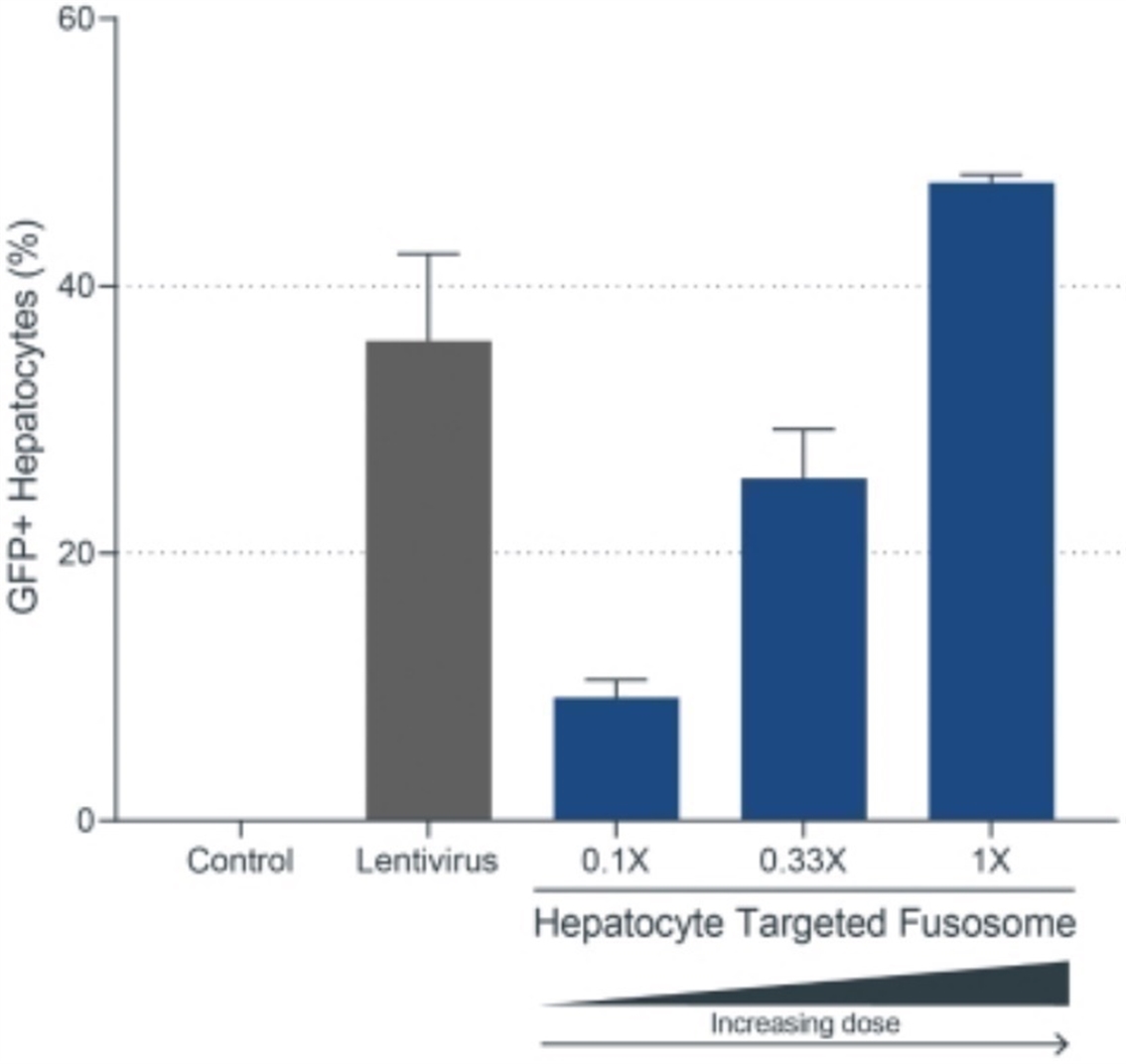
Hepatocyte-targeted fusosomes show levels of in vivo transduction similar to conventional lentivirus and dose dependence. Fusosomes were delivered by intravenous injection (tail vein) into humanized liver mice (FRG, or FAH- immunodeficient mice, repopulated with primary human hepatocytes). Dose for lentivirus was 1.4 1011 TU/kg (# of transducing units per kilogram mouse body weight, tested on primary human hepatocytes). Highest dose for the hepatocyte-targeted fusosome was 1.1 1010 TU/kg (1X) and lower doses were at one-third (0.33X) and one-tenth (0.1X) of the highest dose. Liver cells were harvested from injected mice seven days after injection, separated into hepatocytes and non-parenchymal cells (non-hepatocytes) and analyzed for GFP expression and species (human or mouse).
Background on OTC Deficiency
OTC deficiency is the most common inherited disorder of the urea cycle, the process by which the body detoxifies ammonia and produces urea. It is the only urea cycle disorder that is X-linked, leading to more severe disease in males. OTC deficiency occurs in approximately 1 in 50,000 births, and there are approximately 10,000 patients worldwide. A deficiency of the OTC enzyme leads to accumulation of ammonia, which can lead to neurotoxicity manifesting early as vomiting and anorexia, before progressing to a progressive lethargy, seizures, intellectual impairment, coma, and death. The severity and age of onset of OTC deficiency can vary with the most severely affected, typically males, presenting shortly after birth. In this severe, neonatal onset of OTC deficiency, patients present with an overwhelming illness that rapidly progresses with up to 90% mortality rate despite advances in standard of care treatments. In less severely affected patients who present later in childhood or as adults, severe elevations of ammonia and resulting neurotoxicity still occur, primarily precipitated by an illness or excessive protein intake.
OTC Deficiency: Current Treatment Landscape and Unmet Need
The standard of care for patients with OTC deficiency includes a low protein diet, nutrient supplementation, and the use of ammonia scavengers such as benzoate, phenylacetate, or phenylbutyrate. Despite all of these measures, patients may still experience acute hyperammonemia crises particularly in the setting of increased protein catabolism that can be induced by viral illness or certain medications. These acute crises are treated with supportive care including kidney dialysis for rapid ammonia reduction. The frequency and duration of hyperammonemia crises has been directly linked to poor long-term outcomes and intellectual disability. The only curative therapy available is liver transplantation, which has become more common as surgical techniques and supportive care have improved over time. In those patients with severe, neonatal onset of OTC deficiency, liver transplantation is commonly performed before the age of five and, in some cases, can occur before one year of age.
In addition to the standard of care therapies noted above, therapies to replace the defective OTC gene have been pursued. Recent trials have primarily utilized AAVs to deliver a corrected OTC gene. While these viruses have to date been generally well tolerated, they are still associated with significant immunogenicity that can preclude use in the up to one third of patients with pre-existing antibodies to AAV and can lead to systemic symptoms, including elevated liver enzymes. Beyond the challenge of pre-existing antibodies, the primary drawback is the potential for transient efficacy as the gene replacement via AAV would not be expected to be
54
permanent if replication of the target cell occurs. While the durability of an AAV delivered gene replacement for OTC deficiency depends on many factors, one of the key determinants is the rate of hepatocyte turnover. This is especially relevant in pediatric patients with growing livers and rapid cell turnover. This dilution of effect has been supported by animal studies where AAV delivered gene replacement was successful in adult animals but not successful in younger animals. The most severe form of OTC deficiency presents in the neonatal period and, if a donor is available, may be treated with liver transplantation, a permanent gene therapy that provides long lasting benefit to patients is required to address the greatest unmet need from OTC deficiency. Additionally, as hepatocytes continue to divide approximately once a year even in adults, a durable gene therapy would also be expected to provide an advantage even in adult patients where an AAV delivered gene therapy is likely to lose function over time.
We believe our approach of pursuing a permanent gene replacement therapy has the potential to improve morbidity, mortality, and quality of life even in the youngest, most severely ill patients.
Development Plan and Key Next Steps
We are conducting mouse studies to establish proof of concept and inform the dose profile of our lead hepatocyte fusosome. In the near term, we are seeking to finalize the hepatocyte-targeted fusosome candidate and begin GLP production. Dose and safety of our lead fusosome compositions for OTC will be further informed through NHP studies, and with success, we expect to submit an IND in the next several years. The hepatocytes of the liver are the one cell type readily accessible to competing technologies such as AAVs and LNPs. Each of these technologies has limitations overcome by the fusosome platform, but any future investment in therapeutics targeting hepatocytes will need to consider if these fusosome advantages are meaningful enough to patients to justify the investment. For example, we are currently evaluating whether our technology offers differentiated solutions to these liver-targeted gene therapies for genetic diseases such as Hemophilia and Alpha-1 antitrypsin deficiency.
HSC Fusosome Program
We are developing hematopoietic stem cell (HSC) targeted fusosomes, designed to target and repair genetic abnormalities underlying diseases such as sickle cell disease and beta-thalassemia (SG418), with the goal of achieving preclinical proof of concept as early as 2023.
Background on hemoglobinopathies
Devastating inherited hematologic disorders, including sickle cell disease, beta-thalassemia, and other hemoglobinopathies, are caused by a monogenic variant, and patients suffering from these diseases are candidates for in vivo cell engineering.
Sickle cell disease (SCD) is caused by a single point mutation in the beta globin gene (HbB). The resulting mutant form of the protein, referred to as HbS, is prone to aggregate into long, rigid molecules that deform red blood cells (RBCs) into a sickle shape, obstructing blood vessels and undergoing premature lysis. The consequences are severe pain (sickle cell crisis), tissue infarction, infection, anemia, stroke, and early death. SCD is the most common inherited blood disorder in the United States, affecting an estimated 100,000 individuals, and 134,000 individuals in Europe. The global prevalence of SCD is estimated to be approximately 4.4 million individuals and is most common among people of African, Middle Eastern and South Asian descent.
Beta-thalassemia is an inherited blood disorder caused by any one of over 200 mutations in HbB which results in reduced production of functional hemoglobin. Transfusion-dependent beta-thalassemia (TDBT) is the most severe form of this disease, often requiring multiple transfusions per year. Patients with TDBT suffer from failure to thrive, persistent infections, and life-threatening anemia. Frequent blood transfusions can lead to iron overload that then require iron chelation therapy, which itself is associated with significant toxicities, resulting in low levels of adherence. Even with frequent transfusions, patients with TDBT continue to suffer from failure to thrive, persistent infections, and life-threatening anemia.
The prevalence of beta-thalassemia globally is estimated to be 288,000. The total combined prevalence of beta-thalassemia in the United States and Europe is estimated to be approximately 19,000 patients, mostly in Europe. Of the patients currently treated in the United States and Europe, we believe approximately 50% and 10%, respectively, are transfusion dependent. Beta-thalassemia is especially prevalent in developing countries of Africa, South Asia, Southeast Asia, the Mediterranean region and the Middle East. Although historically prevalent in Mediterranean North Africa and South Asia, thalassemias are now encountered in other regions as a result of changing migration patterns. As such, there is a growing focus on developing new therapeutics aimed at improving quality of life for this significant unmet medical need.
Correction of the causal monogenic defects could potentially provide a one-time, curative treatment approach, rather than the current lifelong, multidisciplinary standard of care treatment.
55
Current Treatment Landscape and Unmet Need
Despite its clear and well-known genetic nature, SCD remains underserved, with existing treatment strategies mostly supportive in nature. Allogeneic HSC transplantation (HSCT) is currently the only potentially curative therapy available. However, HSCT is limited by donor availability (approximately 15-30% worldwide).
Furthermore, chronic graft-versus-host disease is a major risk that contributes to the long-term morbidities associated with allogeneic HSCT. Otherwise, treatment options largely manage disease symptoms, including analgesia during crises, hydroxyurea, L-glutamine, and anti-infectives. Recently, two disease-modifying treatments were approved by the FDA, crizanlizumab and voxelotor. Crizanlizumab was approved for treating crises in SCD patients who are unresponsive to either hydroxyurea or L-glutamine. Voxelotor is an oral small molecule inhibitor of HbS polymerization, which compared to placebo, was associated with a reduction in acute crises. While these agents represent a meaningful advance in the treatment of SCD, they focus on supportive care and do not address the mutation in the gene that is the root cause of the disease.
As in SCD, there are limited treatment options available for TDBT, and those that exist are supportive in nature. Allogeneic HSCT is similarly potentially curative but is also limited by donor availability, the risk of GVHD, and other comorbidities that result from the procedure. Because of the need for recurring blood transfusions, patients require ongoing chelation therapy to avoid iron load from the transfusions and its associated organ damage. However, this treatment is burdensome and associated with significant toxicities, and consequently, has low adherence. Currently, there is only one FDA approved therapy for beta-thalassemia, luspatercept, which significantly reduces the frequency of blood transfusions needed. However, safety concerns remain with a possible increased risk for hypertension and thromboembolic events.
There are several therapies in development to treat diseases of the hematopoietic system that have demonstrated clinical proof of concept through ex vivo gene modification. These approaches directly address the genetic activity missing in SCD and TDBT by supplying a novel gene to the patient’s cell or by editing genome to enhance hemoglobin expression. The ex vivo process begins with the mobilization and removal of cells from the blood, a process known as leukapheresis. Next, these cells undergo a process to enrich for cells expressing an HSC marker, CD34. The enrichment of CD34 cells increases the percentage of long-lived HSCs, the key stem cell that is both persistent and can differentiate into all the cells of the blood. However, even under enrichment, long-lived HSCs make up less than 1% of all the CD34 cells. CD34+ cells are transduced with either a novel gene or genome editing complexes, each having a distinct therapeutic action. The cells are then cryopreserved and sent back to the patient. Before transplantation, the patients receive conditioning chemotherapy to prepare the body so that the gene-modified cells engraft after re-infusion. The current conditioning regimens are toxic, with significant risks and side effects, although less toxic regimens are in development. Key questions remain regarding durability and safety, particularly over time, for transplanting these ex vivo modified HSCs. Furthermore, manufacturing complexities, cost, and the complications from the myeloablative conditioning chemotherapy regimens remain significant obstacles to widespread adoption. There are multiple ongoing efforts to improve this approach by focusing on HSC procurement, transduction, gene-editing, milder conditioning regimens, and transplantation efficiency. We believe that the most meaningful opportunity to improve outcomes is to eliminate the complex ex vivo modification and transplantation steps by utilizing our fusogen technology to develop fusosomes that specifically target HSC and other key hematopoietic cells via in vivo delivery.
Our HSC Fusosome Approach
The use of an in vivo fusosome-based delivery system bypasses the requirement for ex vivo manufacturing and would require no conditioning chemotherapy. Without the manufacturing complexity and the requisite hospital stay for a patient who has undergone conditioning, as well as the concomitant costs and risks of each, in vivo therapies have the potential to meaningfully increase the number of patients that receive these therapies.
Targeting HSCs in vivo using fusogens requires identifying the appropriate cells and their corresponding cell surface receptors. HSCs have no single specific marker, but there are a number of cell surface proteins that are highly enriched on HSCs. Some of these markers also appear on erythrocytic, or red blood cell, progenitors, which may help establish both short-term and long-term efficacy. We have an ongoing program to discover fusogens with appropriate target specificity.
In parallel, we are establishing our capability to deliver different payloads utilizing the fusosome system. Our goal is to establish the appropriate cell specificity with the ability to utilize the appropriate gene modification system to achieve the right outcome for patients. With successful cell-specific targeting, we have an opportunity to deliver the therapeutic payload to the right cell without the need for complex ex vivo manufacturing or toxic conditioning chemotherapy.
56
Development Plan and Key Next Steps
The next major milestones are to identify candidate fusogens for specific HSC targeting and fusosome compositions with relevant genome modification payloads. Our goal is to achieve preclinical proof of concept for SG418 as early as 2023 and submit an IND in the next several years.
SanaX
Despite the significant advances in the development of successful cell and gene therapies that have been made to date, there remain a number of fundamental limitations of existing technologies that prevent achieving the maximal breadth of application of these new therapeutic approaches. We wish to lead both the present and future of cell and gene therapy, and we are therefore committed to investing in research and other activities that will ensure a leadership position for the long-term. Towards this end, we have established SanaX as a distinct research arm.
In contrast to the industry’s traditional research activities which are focused primarily on near-term product development using existing technologies, SanaX is devoted to finding solutions to the limitations of today’s technology in order to expand the breadth of therapeutic opportunities. SanaX research efforts are aimed at making fundamental improvements to existing technologies and establishing new paradigms for gene and cell delivery that will ultimately lead to the development of completely new therapeutic modalities.
Truly novel technology development requires the unique ability to thoughtfully marry rigorous experimental science with specific technical goals. Often, fundamental biological problems must be understood in depth in order to define the pathway to a new technological and therapeutic capability. SanaX has established a unique physical and cultural environment with individuals that possess the requisite intellectual and technical capabilities essential for success. One characteristic of the SanaX research environment that we believe will be extremely valuable is a “nimbleness” that enables the team to immediately embrace new technical or scientific information and/or meet specific unanticipated therapeutic needs. In addition, several collaborative efforts with outside investigators possessing specific biological sector expertise have been established to enhance our internal efforts.
Current SanaX research activities are focused in several areas where we believe advances in technology are most critical. Some of these efforts include:
|
|
• |
evaluating the use of cells, rather than viruses, as delivery vehicles; |
|
|
• |
re-purposing several different virus vector systems and virus-like particles (VLPs) to expand the therapeutic payloads that may be delivered by the different viruses and VLPs; |
|
|
• |
developing novel approaches to the production of different viral vectors; |
|
|
• |
developing novel methods for enabling the exogenous control of transgene expression via small molecule drugs; |
|
|
• |
developing new paradigms for genetically manipulating specific arms of the immune response in order to engender immunological tolerance to specific antigens, cells, and organs; and |
|
|
• |
COVID-19 related research focused on the delivery of specific anti-SARS-Cov-2 antibodies and the evaluation of novel direct anti-viral strategies. |
Dr. Mulligan, our Executive Vice-Chairman and Head of SanaX, directly oversees the SanaX research effort. SanaX maintains an independent research budget in order to ensure that these longer-term, disruptive priorities are not sacrificed for near-term needs. Once SanaX develops an understanding of how a technology can translate into the clinic, a program will move from SanaX into our internal R&D and manufacturing organization or partnered externally.
Manufacturing Strategy and Approach
While the field of cell and gene therapy has had a number of successes with innovative therapies, the challenges of manufacturing at industrial scale have limited access for patients in need. As was the case during the initial development of recombinant biologics, an improvement to our ability to characterize these products will be essential to increasing patient access. It is especially critical to have an in-depth understanding of the impact of manufacturing processes on the product quality attributes and resulting clinical performance of the product.
From inception, we have recognized the key role manufacturing plays in enabling the access of these innovative engineered cells as medicines. Two areas of particular focus are product analytical and biological characterization, leading to a better definition of critical product attributes, as well as process understanding, leading to better control the impact of process parameters on these critical product attributes.
57
We have developed a manufacturing strategy that supports our vision of democratizing access with early investments in people, technology, and infrastructure:
|
|
• |
establishing a team with diverse, experienced talents with extensive knowledge of both the process and analytical sciences in the field of cell and gene therapy, as well as CMC product development expertise from preclinical to global commercialization; |
|
|
• |
establishing multiple manufacturing platforms for our diverse portfolio; and |
|
|
• |
establishing infrastructure from lab bench to a GMP manufacturing and supply chain network. |
To support our ex vivo and in vivo development pipeline, we are initially establishing three manufacturing platforms: viral vector, allogeneic T cells, and PSC-derived.
While the three manufacturing platforms are very different in terms of the manufacturing process and supply chain, they also share some common challenges and opportunities. For example, product characterization and analytical development are critical, and these capabilities are fungible across platforms. In addition, we are focusing on some of the key areas in each of the platforms to enable scaled manufacturing. For the viral vector platform, we are starting early in the research phase with suspension culture process in bioreactors similar to protein biologics to maximize process yield and batch to batch process robustness at scale. Transfer to these bioreactors later in development can complicate product comparability assessments. For the allogeneic T cell program, we are focusing on scaling the multiplex gene editing process and understanding of the impact of the variability of the starting material from healthy donors to on product quality. For stem-cell derived therapies, such as beta cells, cardiomyocytes, and glial progenitor cells, we are focusing on developing a scalable process and analytical technologies to characterize stability of the starting cells, end cell products, and critical product quality attributes.
To establish our manufacturing capability, we started with a non-GMP pilot plant for ex vivo and in vivo engineered cell platform processes with up to 200L bioreactor scale. This provides the infrastructure for process and technology development, technology transfer support, and production for non-GMP material such as GLP toxicology study material. In addition, we are taking a hybrid approach to establish our end-to-end supply chains for the three manufacturing platforms, leveraging a combination of internal manufacturing capability and external contract development and manufacturing organizations (CDMOs) for clinical supplies, in a staged manner:
|
|
• |
we will utilize CDMOs for GMP supplies initially to support our upcoming INDs and early-stage clinical trials; and |
|
|
• |
we intend to build the internal manufacturing facilities needed to support late-stage clinical trials and commercialization of therapies across our pipeline. |
Operating our own internal manufacturing facilities to complement our CDMO networks is a key to our strategy. Accordingly, in July 2021, we entered into a long-term lease to establish and operate our own GMP manufacturing facility to support our late-stage clinical development and early commercial product candidates across our product portfolio, including with the production of allogeneic T cells, viral vectors, and PSC-derived products. We believe that investing in an internal manufacturing facility will offer us a competitive advantage that will better position us to execute on our goal of ensuring broad and uninterrupted patient access to our therapies, including by allowing us to mitigate delays related to third-parties, including related to capacity-, personnel-, or production-related issues at our CDMOs; develop proprietary knowledge and product and process expertise we can utilize across our programs to create long-term value; and design a facility that can be optimized for and adaptable to our existing and future needs.
Competition
There are other companies that have stated that they are developing cell and gene therapies that may address oncology, diabetes, CNS disorders, and cardiovascular diseases. Some of these companies may have substantially greater financial and other resources than we have, such as larger research and development staff and well-established marketing and salesforces, or may operate in jurisdictions where lower standards of evidence are required to bring products to market. For example, we are aware that some of our competitors, including Novartis International AG, Gilead Sciences, Inc., Bristol-Myers Squibb Company, Novo Nordisk A/S, Johnson & Johnson, Allogene Therapeutics, Inc., CRISPR Therapeutics AG, Precision BioSciences, Inc., Caribou Biosciences, Inc., Fate Therapeutics, Inc., Century Therapeutics, Inc., bluebird bio, Inc., 2seventy bio, Inc., Orchard Therapeutics PLC, Aruvant Sciences, Inc., Sanofi S.A., Editas Medicine, Inc., Beam Therapeutics Inc. (Beam), ViaCyte Inc., Vertex Pharmaceuticals Incorporated, Eli Lilly and Company, Astellas Pharma Inc., and Bayer AG might be conducting large-scale clinical trials for therapies that could be competitive with our ex vivo and in vivo programs. Among companies pursuing ex vivo and in vivo cell engineering, we believe we are substantially differentiated by our robust intellectual property portfolio, extensive research, rigorous and objective approach, and multidisciplinary capabilities.
58
Intellectual Property
We strive to protect and enhance the proprietary technology, inventions, and improvements that are commercially important to our business, including seeking, maintaining, and defending patent rights, whether developed internally or licensed from our collaborators or other third parties. Our policy is to seek to protect our proprietary position by, among other methods, filing patent applications in the United States and in jurisdictions outside of the United States related to our proprietary technology, inventions, improvements, and product candidates that are important to the development and implementation of our business. We also rely on trade secrets and know-how relating to our proprietary technology and product candidates, continuing innovation, and in-licensing opportunities to develop, strengthen, and maintain our proprietary position in the field of cell and gene therapy. We additionally plan to rely on data exclusivity, market exclusivity, and patent term extensions when available and, where applicable, plan to seek and rely on regulatory protection afforded through orphan drug designations. Our commercial success may depend in part on our ability to obtain and maintain patent and other proprietary protection for our technology, inventions, and improvements, preserve the confidentiality of our trade secrets, maintain our licenses to use intellectual property owned by third parties, defend and enforce our proprietary rights, including our patents, and operate without infringing on the valid and enforceable patents and other proprietary rights of third parties.
We have in-licensed and developed numerous patents and patent applications, which include claims directed to compositions, methods of use, processes, dosing, and formulations, and possess substantial know-how and trade secrets relating to the development and commercialization of our ex vivo and in vivo cell engineering platforms and related product candidates, including related manufacturing processes. As of February 2022, our in-licensed and owned patent portfolio consisted of approximately 41 licensed U.S. issued patents, approximately 78 licensed U.S. pending patent applications, and approximately 55 owned U.S. pending patent applications, as well as approximately 69 licensed patents issued in jurisdictions outside of the United States, approximately 326 licensed patent applications pending in jurisdictions outside of the United States (including approximately five licensed pending Patent Cooperation Treaty (PCT) applications), and approximately 42 owned patent applications pending in jurisdictions outside of the United States (including approximately 15 owned pending PCT applications) that, in many cases, are counterparts to the foregoing U.S. patents and patent applications. The patents and patent applications outside of the United States in our portfolio are held primarily in Europe, Canada, China, Japan, and Australia. For information related to our in-licensed intellectual property, see the subsection below titled “—Key Intellectual Property Agreements.”
For the product candidates and related manufacturing processes we develop and commercialize in the normal course of business, we intend to pursue, when possible, composition, method of use, process, dosing, and formulation patent protection. We may also pursue patent protection with respect to manufacturing, drug development processes and technology, and our technology platforms. When available to expand our exclusivity, our strategy is to obtain or license additional intellectual property related to current or contemplated development platforms, core elements of technology, and/or product candidates.
Individual patents extend for varying periods of time, depending upon the date of filing of the patent application, the date of patent issuance, and the legal term of patents in the countries in which they are obtained. Generally, patents issued for applications filed in the United States and in many jurisdictions worldwide have a term that extends to 20 years from the earliest nonprovisional filing date. In the United States, a patent’s term may be lengthened by patent term adjustment, which compensates a patentee for administrative delays by the Unites States Patent and Trademark Office (USPTO) in examining and granting a patent counterbalanced by delays on the part of a patentee, or may be shortened if a patent is terminally disclaimed over another patent. In addition, in certain instances, the patent term of a U.S. patent that covers an FDA-approved drug may also be eligible for patent terms extension, which recaptures a portion of the term effectively lost as a result of the testing and regulatory review periods required by FDA. The patent term extension period cannot be longer than five years, and the total patent term, including the extension, cannot exceed 14 years following FDA approval; however, there is no guarantee that the applicable authorities will agree with our assessment of whether such extensions should be granted, and, if granted, the length of such extensions. Similar provisions are available in Europe and other foreign jurisdictions to extend the term of a patent that covers an approved drug. Our patents issued as of February, 2022, have terms expected to expire on dates ranging from 2023 to 2040. If patents are issued on our patent applications pending as of February, 2022, the resulting patents are projected to expire on dates ranging from 2023 to 2043. However, the actual protection afforded by a patent varies on a product-by-product and country-to-country basis and depends upon many factors, including the type of patent, the scope of its coverage, the availability of regulatory-related extensions, the availability of legal remedies in a particular country, and the validity and enforceability of the patent.
In some instances, we submit patent applications directly with the USPTO as provisional patent applications. Provisional applications for patents were designed to provide a lower-cost first patent filing in the United States. Corresponding non-provisional patent applications must be filed not later than 12 months after the provisional application filing date. The corresponding non-provisional application benefits in that the priority date(s) of the patent application is/are the earlier provisional application filing date(s), and the patent term of the finally issued patent is calculated from the later non-provisional application filing date. This system allows us to obtain an early priority date, add material to the patent application(s) during the priority year, obtain a later start to the patent term, and to delay prosecution costs, which may be useful in the event that we decide not to pursue examination in an
59
application. While we intend to timely file nonprovisional patent applications relating to our provisional patent applications, we cannot predict whether any such patent applications will result in the issuance of patents that provide us with any competitive advantage.
We file U.S. nonprovisional applications and PCT applications that claim the benefit of the priority date of earlier filed provisional applications, when applicable. The PCT system allows an applicant to file a single application within 12 months of the original priority date of the patent application and to designate all of the 153 PCT member states in which national patent applications can later be pursued based on the international patent application filed under the PCT. The PCT searching authority performs a patentability search and issues a non-binding patentability opinion which can be used to evaluate the chances of success for the national applications in foreign countries prior to having to incur the filing fees. Although a PCT application does not issue as a patent, it allows the applicant to seek protection in any of the member states through national-phase applications. At the end of the period of two and a half years from the first priority date of the patent application, separate patent applications can be pursued in any of the PCT member states either by direct national filing or in some cases by filing through a regional patent organization, such as the European Patent Organization. The PCT system delays expenses, allows a limited evaluation of the chances of success for national/regional patent applications, and enables substantial savings where applications are abandoned within the first two and a half years of filing.
For all patent applications, we determine claiming strategy on a case-by-case basis. We always consider the advice of counsel and our business model and needs. We file patents containing claims for protection of all useful applications of our proprietary technologies and any product candidates, as well as all new applications and/or uses we discover for existing technologies and product candidates, assuming these are strategically valuable. We continuously reassess the number and type of patent applications, as well as the pending and issued patent claims, to help ensure that maximum coverage and value are obtained for our inventions given existing patent office rules and regulations. Further, claims may be and typically are modified during patent prosecution to meet our intellectual property and business needs.
We recognize that the ability to obtain patent protection and the degree of such protection depends on a number of factors, including the extent of the prior art, the novelty and non-obviousness of the invention, and the ability to satisfy the enablement requirement of the patent laws. In addition, the coverage claimed in a patent application can be significantly reduced before the patent is issued, and its scope can be reinterpreted or further altered even after patent issuance. Consequently, we may not obtain or maintain adequate patent protection for any of our future product candidates or for our technology platform. We cannot predict whether the patent applications we are currently pursuing will issue as patents in any particular jurisdiction or whether the claims of any issued patents will provide sufficient proprietary protection from competitors. Any patents that we hold may be challenged, circumvented, or invalidated by third parties.
The patent positions of companies like ours are generally uncertain and involve complex legal and factual questions. No consistent policy regarding the scope of claims allowable in patents in the fields of cell and gene therapy has emerged in the United States. The patent situation outside of the United States is even more uncertain. Changes in either the patent laws or their interpretation in the United States and worldwide may diminish our ability to protect our inventions and enforce our intellectual property rights, and more generally could affect the value of our intellectual property. In particular, our ability to stop third parties from making, using, selling, offering to sell, or importing products that infringe our intellectual property will depend in part on our success in obtaining and enforcing patent claims that cover our technology, inventions, and improvements. With respect to both licensed and company-owned intellectual property, we cannot be sure that patents will be granted with respect to any of our pending patent applications or with respect to any patent applications filed by us in the future, nor can we be sure that any of our existing patents or any patents that may be granted to us in the future will be commercially useful in protecting our products and the methods used to manufacture those products. Moreover, even our issued patents do not guarantee us the right to practice our technology in relation to the commercialization of our products. The area of patent and other intellectual property rights in biotechnology is an evolving one with many risks and uncertainties, and third parties may have blocking patents that could be used to prevent us from commercializing our patented product candidates and practicing our proprietary technology. It is uncertain whether the issuance of any third-party patent would require us to alter our development or commercial strategies, products, or processes, obtain licenses, or cease certain activities. Our breach of any license agreements or our failure to obtain a license to proprietary rights required to develop or commercialize our future products may have a material adverse impact on us. If third parties prepare and file patent applications in the United States that also claim technology to which we have rights, we may have to participate in interference or derivation proceedings in the USPTO to determine priority of invention. Our issued patents and those that may issue in the future may be challenged, invalidated, or circumvented, which could limit our ability to stop competitors from marketing related products or limit the length of the term of patent protection that we may have for our product candidates. In addition, the rights granted under any issued patents may not provide us with protection or competitive advantages against competitors with similar technology. Furthermore, our competitors may independently develop similar technologies. For these reasons, we may have competition for our product candidates. Moreover, because of the extensive time required for development, testing, and regulatory review of a potential product candidate, it is possible that, before any particular product candidate can be commercialized, any related patent may expire or remain in force for only a short period following commercialization, thereby reducing any advantage of the patent. Our commercial success will also depend in part on not infringing upon the proprietary rights of third parties. Patent disputes are sometimes interwoven into other business disputes.
60
As of February, 2022, our registered trademark portfolio contained approximately 25 registered trademarks and pending trademark applications, consisting of approximately two pending trademark applications in the United States, and approximately 13 registered trademarks and approximately ten pending trademark applications in the following countries through both national filings and under the Madrid Protocol: Australia, Canada, China, European Union, India, Japan, Republic of Korea, Singapore, and Switzerland.
We may also rely, in some circumstances, on confidential information, including trade secrets, to protect our technology. However, trade secrets are difficult to protect. We seek to protect our technology and product candidates, in part, by entering into confidentiality agreements with those who have access to our confidential information, including our employees, contractors, consultants, collaborators, and advisors. We also seek to preserve the integrity and confidentiality of our proprietary technology and processes by maintaining physical security of our premises and physical and electronic security of our information technology systems. Although we have confidence in these individuals, organizations, and systems, agreements or security measures may be breached, and we may not have adequate remedies for any breach. In addition, our trade secrets may otherwise become known or may be independently discovered by competitors. To the extent that our employees, contractors, consultants, collaborators, and advisors use intellectual property owned by others in their work for us, disputes may arise as to the rights in related or resulting know-how and inventions. For this and more comprehensive risks related to our proprietary technology, inventions, improvements, and products, see the subsection titled “Risk Factors —Risks Related to Intellectual Property and Information Technology.”
Key Intellectual Property Agreements
The following describes the key agreements by which we have acquired and maintained certain technology related to our ex vivo and in vivo cell engineering platforms and therapeutic programs.
Ex Vivo Cell Engineering Platform
License Agreement with Harvard
In March 2019, we entered into a license agreement (as amended, the Harvard Agreement) with the President and Fellows of Harvard College (Harvard), pursuant to which we obtained an exclusive, worldwide, sub-licensable license under certain patent rights controlled by Harvard to make, have made, use, offer for sale, sell, have sold and import (i) products and services covered by the patent rights and (ii) products containing stem cells, pluripotent cells or cells derived from stem cells, or pluripotent cells with certain specified genetic modifications ((i) and (ii) together, Harvard Products) or otherwise practice under and exploit the licensed patent rights, for the treatment of disease in humans or, in the case of certain other patent rights, for applications that involve the use of cells derived ex vivo from stem cells in the treatment of disease in humans. We also obtained a non-exclusive, sub-licensable license under certain other patent rights in the United States, and a non-exclusive, sub-licensable, worldwide license under know-how pertaining to the licensed patent rights, to make, have made, use, offer for sale, sell, have sold and import the Harvard Products, or otherwise practice under and exploit the licensed patent rights and know-how, for the treatment of disease in humans. We have the option to obtain such non-exclusive rights in additional jurisdictions if Harvard is successful in obtaining the right to grant such from the third-party co-owner of such patent rights. In October 2021, we entered into an amendment to the Harvard Agreement to include products containing primary cells with certain specified genetic modifications as Harvard Products. We utilize these license rights in our ex vivo cell engineering program relying on our hypoimmune technology.
We are obligated to use commercially reasonable efforts to develop Harvard Products in accordance with a written development plan, to market the Harvard Products following receipt of regulatory approval, and to achieve certain specified development and regulatory milestones within specified time periods, as such period may be extended, for at least two Harvard Products.
The licenses granted pursuant to the Harvard Agreement are subject to certain rights retained by Harvard and the rights of the U.S. government. The retained rights of Harvard pertain only to the ability of Harvard and other not-for-profit research organizations to conduct academic research and educational and scholarly activities and do not limit our ability to pursue our programs and product candidates. We agreed that we will not use any of the licensed patent rights for human germline modification, including intentionally modifying the DNA of human embryos or human reproductive cells.
Pursuant to the Harvard Agreement, we paid Harvard an upfront fee of $3.0 million, and we issued 2.2 million shares of our Series A-2 convertible preferred stock to Harvard as partial consideration for the licenses granted under the Harvard Agreement. Additionally, we paid $6.0 million to Harvard in connection with the issuance of shares of our Series B convertible preferred stock. We paid Harvard annual license maintenance fees of $25,000 for 2019, $50,000 for 2020, and $100,000 for each of 2021 and 2022, and we are required to pay annual license maintenance fees of $100,000 for each calendar year thereafter for the remainder of the term. We are required to pay Harvard up to an aggregate of $15.2 million per Harvard Product upon the achievement of certain pre-specified development and regulatory milestones for up to a total of five Harvard Products, or an aggregate total of $76.0 million for all five Harvard Products. These milestone payments would double if we undergo a change of control. We are also obligated to pay,
61
on a product-by-product and country-by-country basis, royalties in the low single-digit percentage range on quarterly net sales of Harvard Products covered by licensed patent rights, and a lower single-digit percentage royalty on quarterly net sales of Harvard Products not covered by licensed patent rights. The royalty rates with respect to Harvard Products covered by licensed patent rights are also subject to specified and capped reductions for loss of market exclusivity and for payments owed to third parties with respect to patent rights which cover Harvard Products in the territory. We are also obligated to pay Harvard a percentage of certain sublicense income ranging from the high single-digit to low double-digit percentage range. We are obligated to pay up to $175.0 million in success payments based on increases in the per share fair value of our common stock at pre-specified valuation dates that include the one-year anniversary of the consummation of our IPO and periodically thereafter, the date of the consummation of a merger, an asset sale, or merger, or the sale of the majority of the shares held by our Series A convertible preferred stockholders, and the last day of the term of the success payments.
The Harvard Agreement will expire upon the expiration of the last-to-expire valid claim within the licensed patent rights or, if later, at the end of the final royalty term, which is determined on a Harvard Product-by- Harvard Product and country-by-country basis, and is the later of (i) the date on which the last valid claim within the licensed patent rights covering such Harvard Product in such country expires, (ii) expiry of regulatory exclusivity for such Harvard Product in such country, or (iii) ten years from the first commercial sale of such Harvard Product in such country, which we expect to occur in 2039. We also have the right to terminate the Harvard Agreement in its entirety for any reason upon 45 days’ prior written notice to Harvard. Either party may terminate the Harvard Agreement upon a material breach by the other party that is not cured within 60 days after receiving written notice thereof. Harvard may terminate the Harvard Agreement upon written notice in the event of our bankruptcy, insolvency, or similar proceedings. If we terminate the Harvard Agreement for convenience, our obligations to pay milestones and royalties with respect to Harvard Products that are not then covered by licensed patent rights will survive for the remainder for the applicable royalty term. If the Harvard Agreement is terminated for any reason, then sublicensees, other than our affiliates or sublicensees in material default or at fault for the termination, have the right to enter into a direct license with Harvard on substantially the same non-economic terms and on economic terms providing for the payment to Harvard of the consideration that would otherwise have been payable if the Harvard Agreement and the sublicense were not terminated.
License Agreement with UCSF
In January 2019, we entered into a license agreement (as amended, the UCSF Agreement) with The Regents of the University of California (The Regents) acting through its Office of Technology Management, University of California San Francisco (UCSF), pursuant to which we obtained an exclusive license to inventions related to immunoengineered pluripotent cells and derivatives claimed in U.S. and international patents and patent applications (UCSF Patent Rights) by The Regents. The license is to make, have made, use, sell, offer for sale and import licensed products that are covered by such UCSF Patent Rights, provide licensed services, practice licensed methods, and otherwise practice under the UCSF Patent Rights, for use in humans only, in the United States and other countries where The Regents is not prohibited by applicable law from granting such UCSF Patent Rights. We have the right to sublicense our rights granted under the UCSF Agreement to third parties subject to certain terms and conditions. We utilize these license rights in our ex vivo cell engineering platform program that relies on our hypoimmune technology.
We are obligated, directly or through affiliates or sub-licensees, to use commercially reasonable efforts to develop, manufacture, and sell one or more licensed products and licensed services and to bring one or more licensed products or licensed services to market. We are required to use commercially reasonable efforts to obtain all necessary governmental approvals in each country where licensed products or licensed services are manufactured, used, sold, offered for sale, or imported. We are required to spend at least $30.0 million towards research, development, and commercialization of licensed products within five years after the closing of our Series A-2 convertible preferred stock financing. In addition, we are required to achieve certain specified development and regulatory milestones within specified time periods. We have the ability to extend the time periods for achievement of development and regulatory milestones under certain terms set forth in the UCSF Agreement, including payment of extension fees. If we are unable to complete any of the specified milestones by the completion date, or extended completion date, for such milestone, then The Regents has the right and option to either terminate the Agreement, subject to our ability to cure the applicable breach, or convert our exclusive license to a non-exclusive license.
The Regents reserves and retains the right to make, use and practice the invention, and any related technology, and to make and use any products and to practice any process that is the subject of the UCSF Patent Rights (and to grant any of the foregoing rights to other educational and non-profit institutions) for educational and non-commercial research purposes, including publications and other communication of research results. This does not limit our ability to pursue our programs and product candidates.
Pursuant to the UCSF Agreement, we paid an upfront license fee of $100,000 to The Regents, and we issued The Regents 0.7 million shares of our Series A-2 convertible preferred stock. In addition, we entered into an amendment to the UCSF Agreement in December 2020, pursuant to which we issued 37,500 shares of our common stock to The Regents. We are required to pay license maintenance fees ranging from $10,000 on the first anniversary of the date of the UCSF Agreement to $40,000 on the sixth anniversary and continuing annually thereafter. This fee shall not be due if we are selling or exploiting licensed products or licensed
62
services and paying an earned royalty to The Regents on net sales of such licensed products or licensed service. We are required to pay The Regents up to an aggregate of $2.45 million per licensed product upon the achievement of certain pre-specified development and regulatory milestones for the first five licensed products and half such amount for the second five licensed products, for an aggregate total of $18.4 million in development and regulatory milestone payments. Additionally, we are required to pay The Regents up to an aggregate of $0.5 million per licensed product upon the achievement of certain commercial milestones for the first five licensed products and half such amount for the second five licensed products, for an aggregate total of $3.75 million in commercial milestone payments. With respect to each licensed product, licensed service, or licensed method, we are obligated to pay, on a country-by-country basis, tiered royalties on net sales in the low single-digits. The royalty rates are subject to specified capped reductions for payments owed to unaffiliated third parties in consideration for patent rights, or patent rights together with know-how, in order to practice licensed methods or to make, have made, use sell, offer to sell, or import licensed products or licensed services. We are required to pay to The Regents a minimum annual royalty of $100,000 beginning with the year of the first sale of a licensed product or licensed service and ending upon the expiration of the last-to-expire UCSF Patent Right. This will be credited against any earned royalty due for the twelve-month period following which the minimum payment was made and pro-rated. We are also obligated to pay The Regents a percentage of certain non-royalty sublicense income ranging from the low double-digits to mid-twenties.
The UCSF Agreement will expire on expiration or abandonment of the last valid claims within the UCSF Patent Rights licensed thereunder, which we expect to occur in 2040. The Regents has the right to terminate the Agreement if we fail to cure or discontinue a material breach within 60 days of receiving a notice of default. We have the right to terminate the UCSF Agreement in its entirety or under certain UCSF Patent Rights on a country-by-country basis at any time by providing 60 days’ notice of termination to The Regents. The UCSF Agreement will automatically terminate in the event of our bankruptcy that is not dismissed within a specified time period. The Regents may immediately terminate the Agreement upon written notice if we file a non-defensive patent challenge. The termination of the UCSF Agreement will not relieve us of obligations to pay any fees, royalties, or other payments owed to The Regents at the time of such termination or expiration, including the right to receive earned royalties. If the UCSF Agreement is terminated for any reason, then, upon the request of any sublicensee, The Regents will enter into a direct license with such sublicensee on the same terms as the UCSF Agreement, taking into account any difference in license scope, territory, and duration of sublicense grant, provided that such sublicensee is not at the time of such termination in breach of its sublicensing agreement and is not at the time of such termination an opposing party in any legal proceeding against The Regents.
2019 Exclusive License Agreement with Washington University
In November 2019, we entered into a license agreement (the 2019 WU Agreement) with Washington University, pursuant to which we obtained an exclusive sublicensable, non-transferable, worldwide license under certain Washington University patent rights related to genetically engineered hypoimmunogenic stem cells to research, develop, make, have made, and sell products, the manufacture, use, sale or import of which by us or our sublicensees would, in the absence of the 2019 WU Agreement, infringe at least one valid claim of the licensed patent rights (WU Hypoimmune Products).
We are obligated to use commercially reasonable efforts to (i) develop, manufacture, promote and sell WU Hypoimmune Products and (ii) to achieve certain development, regulatory, and commercial diligence milestones within specified time periods. We have the ability to extend the time periods for achievement of such milestones under certain terms set forth in the 2019 WU Agreement, including payment of extension fees.
Washington University retains the right to make, have made, use, and import WU Hypoimmune Products in fields relating to diagnosis, prevention, and treatment of human diseases or disorders for research and educational purposes, including collaboration with other nonprofit entities, but excluding any commercial purposes, and such retained rights do not limit our ability to pursue our programs and product candidates. Washington University retains all rights not granted to us under the patents. In addition, the 2019 WU Agreement is subject to certain rights retained by the U.S. government, including the requirement that licensed products sold in the U.S. be substantially manufactured in the U.S.
Pursuant to the 2019 WU Agreement, we paid Washington University an upfront fee of $75,000. We are required to pay Washington University up to $100,000 in license maintenance fees on each anniversary of the 2019 WU Agreement’s effective date until the first commercial sale of a WU Hypoimmune Product. Upon the achievement of certain development and regulatory milestones, we are required to pay Washington University up to an aggregate of $2.0 million in milestone payments per WU Hypoimmune Product for the first three WU Hypoimmune Products, for an aggregate of $6 million in development and regulatory milestones. Additionally, upon the achievement of certain commercial milestones, we are required to pay Washington University up to an aggregate of $2.5 million in milestone payments per WU Hypoimmune Product for the first three WU Hypoimmune Products, for an aggregate of $7.5 million in commercial milestones. We are also obligated to pay royalties on annual net sales in the low single-digits, subject to a minimum amount of royalties payable in advance. The minimum annual royalty for the first anniversary of the effective date following the first commercial sale will be $100,000 and subsequently will increase up to a maximum minimum annual royalty of $750,000 on the fourth anniversary of the effective date following the first commercial sale. The royalties are payable
63
provided there is at least one valid claim of licensed patent rights present in the country of manufacture or sale. The royalty rates are also subject to specified and capped reduction upon certain other events. Furthermore, we are obligated to pay Washington University a percentage of certain non-royalty sublicense income in the low double-digits.
The 2019 WU Agreement will expire upon the last-to-expire valid claim in the licensed patent rights, which we expect to occur in 2038. We also have the right to terminate the 2019 WU Agreement for any reason upon 90 days’ prior written notice to Washington University. Washington University may terminate the 2019 WU Agreement upon our material breach that is not cured within 30 days after receiving written notice thereof. In addition, Washington University may terminate the 2019 WU Agreement (i) upon 30 days’ written notice if we fail to achieve certain development, regulatory, or commercial diligence milestones and are unable to resolve Washington University’s concerns through good faith negotiations in accordance with the 2019 WU Agreement, (ii) upon our bankruptcy or insolvency, or (iii) if an order is made or a notice is issued convening a meeting of our stockholders to consider the passing of a resolution of our winding up or a resolution is passed for our winding up (in each case, other than for the purpose of amalgamation or reconstruction). If the 2019 WU Agreement terminates prior to the expiration of the last-to-expire licensed patent rights, we agree (i) to promptly discontinue the exportation of licensed products, (ii) to promptly discontinue the manufacture, sale, and distribution of the licensed products, (iii) to promptly destroy all licensed products in inventory, and (iv) not to manufacture, sell, or distribute licensed products until the expiration of the applicable last-to-expire licensed patent rights.
2020 License Agreement with Washington University
In September 2020, we entered into an exclusive license agreement (the 2020 WU Agreement) with Washington University for certain patent rights relating to the methods and compositions of generating cells of endodermal lineage and beta cells and uses thereof. Under the 2020 WU Agreement, we obtained an exclusive, worldwide, non-transferable, and royalty-bearing license under the patent rights to research, develop, make, have made, sell, offer for sale, have sold, use, have used, export, and import licensed products, the manufacture, use, sale or import of which by us or our sublicensees would, in the absence of the 2020 WU Agreement, infringe at least one valid claim of the licensed patent rights, solely in fields relating to diagnosis, prevention, and treatment of human diseases or disorders. We utilize these license rights in our ex vivo cell engineering platform program that relies on our hypoimmune technology, including our beta cell program.
Under the 2020 WU Agreement, we are obligated to use commercially reasonable efforts to (i) develop, manufacture, promote, and sell licensed products, and (ii) achieve certain development, regulatory, and commercial diligence milestones within specified time periods. We have the ability to extend the time periods for achievement of such milestones under certain terms set forth in the 2020 WU Agreement, including payment of extension fees.
Washington University retains the right to use the licensed patent rights to make, have made, use, and import licensed products worldwide in fields relating to diagnosis, prevention, and treatment of human disease or disorders for research and educational purposes, including collaboration with other nonprofit entities, but expressly excluding any commercial purposes, and such retained rights do not limit our ability to pursue our programs and product candidates. In addition, the 2020 WU Agreement is subject to certain rights retained by the U.S. government, including the requirement that licensed products sold in the U.S. be substantially manufactured in the U.S.
Pursuant to the 2020 WU Agreement, we paid Washington University an upfront license issue fee of $150,000. We are required to pay annual license maintenance fees on each anniversary of the 2020 WU Agreement’s effective date until the first commercial sale of a licensed product. The license maintenance fee for the first and second anniversaries of the effective date will be $25,000 and subsequently will increase by $25,000 per two anniversaries up to a maximum annual license maintenance fee of $100,000. We are also required to pay Washington University up to an aggregate of $2.0 million upon the achievement of certain pre-specified development and regulatory milestones per licensed product for the first three licensed products under the 2020 WU Agreement, for an aggregate of $6 million in development and regulatory milestones. Additionally, we are required to pay Washington University, up to an aggregate of $4.5 million upon the achievement of certain pre-specified commercial milestones per licensed product for the first three licensed products under the 2020 WU Agreement, for an aggregate of $13.5 million in commercial milestones. We are also required to pay, for each licensed product made or sold by or for us worldwide, earned royalties on net sales of the licensed products in the low single-digits, with the royalty rate being subject to specified and capped reduction upon certain events. Under the 2020 WU Agreement, we are obligated to pay a minimum annual royalty commencing with the first anniversary of the effective date following the first commercial sale of the licensed product, which will be paid as an advance against the earned royalties paid to Washington University over the ensuing 12-month period. The minimum annual royalty for the first anniversary of the effective date following the first commercial sale will be $100,000 and subsequently will increase up to a maximum minimum annual royalty of $750,000 on the fourth anniversary of the effective date following the first commercial sale. The royalties are payable provided there is at least one valid claim of the licensed patent rights present in the country of manufacture or sale. Furthermore, we are obligated to pay Washington University a percentage of certain non-royalty sublicense income in the low double-digits.
64
The 2020 WU Agreement will expire upon the last-to-expire valid claim under the licensed patent rights, which we expect to occur in 2038. We also have the right to terminate the 2020 WU Agreement for any reason upon 90 days’ prior written notice to Washington University. Washington University may terminate the 2020 WU Agreement upon our material breach that is not cured within 30 days after receiving written notice thereof. In addition, Washington University may terminate the 2020 WU Agreement (i) upon 30 days’ written notice if we fail to achieve certain development, regulatory, or commercial diligence milestones and are unable to resolve Washington University’s concerns through good faith negotiations in accordance with the 2020 WU Agreement, (ii) upon our bankruptcy or insolvency, or (iii) if an order is made or a notice is issued convening a meeting of our stockholders to consider the passing of a resolution of our winding up or a resolution is passed for our winding up (in each case, other than for the purpose of amalgamation or reconstruction). If the 2020 WU Agreement terminates prior to the expiration of the last-to-expire licensed patent rights, we agree (i) to promptly discontinue the exportation of licensed products, (ii) to promptly discontinue the manufacture, sale and distribution of the licensed products, (iii) to promptly destroy all licensed products in inventory, and (iv) not to manufacture, sell, or distribute licensed products until the expiration of the applicable last-to-expire licensed patent rights.
Oscine Acquisition
In September 2020, we acquired Oscine Corp. (Oscine), a privately-held early-stage biotechnology company pursing a glial progenitor ex vivo cell engineering program, in exchange for $8.5 million in cash, net of certain expenses. Of the total purchase price, $7.6 million was an upfront cash payment and $0.9 million was set aside (the Oscine Holdback Amount) to satisfy certain general representations and warranties as set forth in the stock purchase agreement. We had originally entered into a collaboration, license and option to purchase agreement with Oscine in November 2018. That agreement was terminated upon the closing of our acquisition of Oscine. As part of the Oscine acquisition, we also agreed to pay additional amounts of up to an aggregate of $225.8 million upon achievement of certain pre-specified development and commercial milestones, which we may pay in cash or in shares of our common stock, subject to certain conditions. As a result of the Oscine acquisition, we entered into, or obtained and amended, licenses to various technologies related to our glial progenitor ex vivo cell-based therapy program, including a license agreement with University of Rochester and a seed bank supply agreement with Hadasit Medical Research Services and Development Ltd.
License Agreement with University of Rochester
Effective as of the closing of the Oscine acquisition, we entered into an amended and restated exclusive license agreement (the Rochester Agreement) with the University of Rochester, which amended and restated a prior license agreement between Oscine and its affiliates and the University of Rochester and assigned Oscine’s rights and obligations in the license agreement to us. Under the Rochester Agreement, we obtained an exclusive, royalty-bearing, sublicensable, worldwide license under certain patents, and a non-exclusive, royalty-free license under know-how, to research, develop, import, make, have made, use, sell, offer to sell, commercialize, and otherwise exploit cell-based therapies for the treatment of human central nervous system disease and disorders. We utilize these license rights in our glial progenitor cell-based therapy program. We granted the University of Rochester a license to practice any patent rights that cover inventions in the field of cell-based therapies for human central nervous system diseases and disorders, which inventions are first conceived and reduced to practice solely by Dr. Steven Goldman acting in his capacity as our employee, or jointly with any of our employees reporting to Dr. Goldman, solely for Dr. Goldman or any of his lab members at the University of Rochester to practice such patent rights within Dr. Goldman’s laboratory at the University of Rochester for internal academic research purposes. University of Rochester granted us an automatic royalty-free non-exclusive license, and the option to obtain exclusive rights, to any patent rights or inventions conceived or reduced to practice by Dr. Goldman or members of his laboratory at the University of Rochester within a certain timeframe in connection with the internal academic research license that we granted to the University of Rochester. We are obligated to use commercially reasonable efforts to proceed with the commercial exploitation of the patents, to create a reasonable supply of licensed products to meet demand, and to adhere to a specified commercial development plan for development of stem cell therapy products, with pre-specified development milestones, including obtaining government approvals to market at least one licensed product, and to market such product within twelve months of receiving such approval.
The licenses granted pursuant to the Rochester Agreement are subject to certain rights retained by the University of Rochester and the rights of the U.S. government. The retained rights of the University of Rochester pertain only to its ability to conduct internal academic research other than clinical research and for teaching, education, and other non-commercial research activities, in publications related to its scientific research and findings, and for any other non-clinical and non-commercial purpose that is not inconsistent with the rights granted to us under the Rochester Agreement. These retained rights do not limit our ability to pursue our programs and product candidates.
Pursuant to the Rochester Agreement, we are obligated to pay to University of Rochester minimum annual royalties beginning in January 2023, the amount of which payments will be $20,000 in 2023, $50,000 in 2025, and will increase to $70,000 in 2028 and beyond. The minimum annual royalty payment is creditable against our obligation to pay tiered royalties on annual net sales in the low single-digits. The royalty rates are also subject to reduction upon certain other events. We are also required to pay University of Rochester up to an aggregate of $950,000 upon the achievement of certain pre-specified development and commercial milestones for
65
each licensed product. In addition, we are required to pay a tiered mid-single digit to mid-double digit percentage of revenue arising from any sublicenses granted by us to third parties.
The Rochester Agreement will terminate on the last-to-expire of the licensed patents thereunder, which we expect to occur in 2038. We also have the right to terminate the Rochester Agreement in its entirety for any reason upon 90 days’ prior written notice to the University of Rochester. The University of Rochester may terminate the Rochester Agreement upon our material breach that is not cured within 30 days of receiving written notice thereof or immediately in the event of our bankruptcy. The University of Rochester may also terminate the Rochester Agreement, or at its sole discretion terminate the exclusivity of the license granted, upon our failure to meet the diligence obligations under and cure such failure within 90 days of our receipt of notice thereof, or such longer reasonable time determined by University of Rochester, at its discretion, and subject to a good faith negotiation mechanism included in the Rochester Agreement.
Supply Agreement with Hadasit Medical Research Services and Development Ltd.
In July 2018, Oscine Therapeutics (U.S.) Inc., an affiliate of Oscine, entered into a supply agreement (as amended, the Hadasit Agreement) with Hadasit Medical Research Services and Development Ltd. (Hadasit), pursuant to which Oscine obtained a quantity of seed bank cells and accompanying regulatory information on a non-exclusive basis for the sole purpose of developing, manufacturing, and selling cell therapy products for the treatment or prevention of central nervous system disorders in humans, which cell therapy products are derived using the Oscine proprietary differentiation technology from a certain human ESC line provided by Hadasit under the Hadasit Agreement. We utilize these cells and information in our glial progenitor cell program. Concurrently with our acquisition of Oscine in September 2020, the Hadasit Agreement was assigned by Oscine Therapeutics (U.S.) Inc. to Oscine. We amended the Hadasit Agreement effective as of the closing of the Oscine acquisition, and we subsequently assigned the Hadasit Agreement from Oscine to us.
Pursuant to the Hadasit Agreement, Oscine Therapeutics (U.S.) Inc. paid Hadasit an upfront fee of $24,000. We are required to pay Hadasit up to an aggregate of $1.1 million upon the achievement of certain development milestones for the first product. We are also obligated to pay tiered royalties in the low single digits on annual net sales of the relevant products worldwide, which obligation shall commence upon the first commercial sale of a relevant product and shall expire after 15 years on a product-by-product and country-by-country basis. The royalty rates are also subject to reduction upon certain other events.
The Hadasit Agreement will continue until terminated in accordance with its terms. Hadasit may terminate the Hadasit Agreement upon giving 30 days’ written notice if we fail to make any payment due thereunder and do not cure such failure within 30 days’ notice, or upon 60 days’ written notice if we cease to use the seed bank cells for the development and manufacture of our products, subject to our ability to dispute Hadasit’s claim and resolution of such dispute in accordance with a process set forth in the Hadasit Agreement. Either party may terminate the Hadasit Agreement upon a material breach by the other party that is not cured within 60 days after receiving written notice thereof, or upon giving written notice thereof, in the event of the other party’s bankruptcy.
Cytocardia Acquisition
In November 2019, we acquired Cytocardia, Inc. (Cytocardia), a privately-held early-stage biotechnology company developing ex vivo cell engineering programs focused on the replacement of damaged heart cells, in exchange for $8.0 million in cash, net of certain indebtedness and expenses, of which $6.8 million was an upfront cash payment, and $1.2 million was set aside (Cytocardia Holdback Amount) to satisfy certain general representations and warranties set forth in the stock purchase agreement. We also agreed to pay additional amounts of up to an aggregate of $75.0 million upon our achievement of certain pre-specified development milestones and up to an aggregate of $65.0 million in pre-specified commercial milestones. As a result of that transaction, we obtained licenses to various intellectual property and technologies, including intellectual property and technology related to our cardiomyocyte program that we rely on for development of our cardiac cell therapy product candidates. These included a license agreement with the University of Washington, as described below.
University of Washington
In October 2018, Cytocardia entered into an exclusive start-up license agreement (as amended, the UW Agreement) with the University of Washington (UW), pursuant to which Cytocardia obtained an exclusive license under certain patents relating to stem cell-derived cardiomyocytes and heart regeneration owned solely by UW or jointly by UW and the University of Cambridge, for which UW has the sole right to control the protection and licensing pursuant an inter-institutional agreement between UW and the University of Cambridge. We amended the UW Agreement in November 2019, concurrently with the closing of our acquisition of Cytocardia, and in July 2020 assigned the UW Agreement from Cytocardia to us. We further amended the UW Agreement in January 2021, February 2021, March 2021, April 2021, July 2021, September 2021, and October 2021 to add additional patent families to the scope of the license. The scope of the license is to make, have made, use, offer to sell, sell, offer to lease or lease, import, or otherwise
66
offer to dispose of products worldwide (i) for any use, with respect to certain specified licensed patents, (ii) for the production of cardiomyocytes having an atrial or ventricular phenotype, with respect to other specified licensed patents, and (iii) heart regeneration therapy, with respect to other specified licensed patents. Additionally, UW granted us a non-exclusive, worldwide license to use certain related know-how, clinical trial information, and program materials. We may sublicense the exclusively licensed rights under the UW Agreement. We may also sublicense the non-exclusively licensed rights, but only for the purpose of using them in conjunction with exclusively licensed rights. We utilize intellectual property in our cardiomyocyte program. For a period of 12 months after the effective date of the UW Agreement, UW agreed to provide reasonable written notice to us of any improvements to the licensed patents upon notice to UW.
We have the option to add such improvements to the licensed patents. Pursuant to the UW Agreement, we are required to use commercially reasonable efforts to commercialize the licensed rights and to make and sell licensed products as soon as practicable and to maximize sales thereof. We are also obligated to achieve specified development, regulatory, and commercial milestones within specified time periods.
Inventions covered in the licensed patents have arisen, in whole or in part, from federally supported research by the U.S. federal government, and the licenses granted pursuant to the UW Agreement are subject to certain rights of the U.S. government. UW has retained for itself as well as for Cambridge University and for any other not-for-profit academic research institution, an irrevocable, nonexclusive right to practice the licensed rights for academic research and instructional or any other academic or non-commercial purpose. UW has retained for itself an irrevocable, nonexclusive license to practice licensed rights for clinical purposes. Cambridge University has also retained for itself an irrevocable, nonexclusive license to practice certain rights co-owned with UW for clinical purposes.
Pursuant to the UW Agreement, we will pay to UW a low single-digit royalty on net sales of products, with the royalty rate being subject to specified and capped reduction upon certain events. We will pay minimum annual fees for the term of the UW Agreement, to be creditable against running royalty payments for the preceding calendar year on a noncumulative basis. These minimum annual fees are due following the second anniversary of the effective date of the UW Agreement and continue during the term of the UW Agreement, ranging from $5,000 up to $50,000 for the years following the second anniversary of the first commercial sale of an FDA-approved licensed product. We will also pay to UW non-cumulative, non-creditable, and non-refundable development milestone payments of up to $175,000 and commercial milestone payments of up to $700,000 for the first licensed product to achieve each applicable milestone event. Furthermore, pursuant to the UW Agreement, we are obligated to pay UW a percentage of certain non-royalty sublicense income ranging from the low single-digits to middle double-digits, depending on the stage of development of our licensed products at the time of execution of the sublicense agreement.
The UW Agreement will expire, without further action by the parties, when all valid claims of the licensed patents have expired, and we have sold all licensed products manufactured prior to the expiration of such valid claims, which we expect to occur in 2040. UW may terminate the UW Agreement if we (i) permanently cease operations, (ii) voluntarily file or have filed against us a petition under applicable bankruptcy or insolvency laws that we fail to have released within 30 days after filing, (iii) propose any dissolution, composition, or financial reorganization with creditors, or if a receiver, trustee, custodian, or similar agent is appointed, (iv) make a general assignment for the benefit of creditors, (v) challenge the validity of the licensed patents, or (vi) breach our material obligations under the UW Agreement and do not cure such breach within 60 days. We may terminate the UW Agreement at any time by delivering to UW a written notice of termination at least 60 days prior to the effective date of termination. In addition, we may propose to terminate certain of our licensed rights under the UW Agreement by delivering to UW a written notice of termination accompanied by a proposed written amendment to the UW Agreement at least 60 days prior to the effective date of termination of such licensed rights.
Non-Exclusive License and Development Agreement with FUJIFILM Cellular Dynamics, Inc.
In February 2021, we entered into a non-exclusive license and development agreement (as amended, the FCDI Agreement) with FUJIFILM Cellular Dynamics, Inc. (FCDI), pursuant to which we obtained non-exclusive rights and a license under certain intellectual property rights controlled by FCDI (including intellectual property rights owned by FCDI and patent rights in-licensed from the Wisconsin Alumni Research Foundation) to research, develop, make, have made, use, have used, sell, offer for sale, import, and otherwise exploit human cell therapy products derived from certain iPSC lines for the treatment or prevention of certain diseases. We anticipate utilizing these intellectual property rights and iPSC lines in certain of our ex vivo cell engineering programs.
Pursuant to the FCDI Agreement, we agreed to pay FCDI an upfront fee of $1.0 million, annual license maintenance fees, and license fees of up to $500,000 per indication for one certain cell type or up to $350,000 per indication for certain other cell types. We are required to pay FCDI up to an aggregate of $28.5 million per indication upon the achievement of certain pre-specified development and regulatory milestones for up to a total of three indications and up to an aggregate of $14.25 million in pre-specified development and regulatory milestones for each additional indication. We are also required to pay up to an aggregate of $8.8 million per product upon the achievement of certain pre-specified commercial milestones. In addition, we are obligated to pay royalties on
67
annual net sales of the relevant products worldwide in the low- to mid-single digits, which obligation shall commence upon the first commercial sale of a relevant product and shall expire after 15 years on a product-by-product and country-by-country basis. The royalty rates are also subject to reduction upon certain other events.
The FCDI Agreement will continue until terminated in accordance with its terms. FCDI may terminate the FCDI Agreement upon giving written notice if we fail to make any payment due or upon our material breach, subject, in each case, to our ability to dispute or cure such breach. We may also terminate the FCDI Agreement for convenience upon prior written notice, and either party may terminate upon giving written notice in the event of the other party’s bankruptcy.
License Agreement with Beam
In October 2021, we entered into an option and license agreement (the Beam Agreement) with Beam, pursuant to which Beam granted us a non-exclusive license to use Beam’s proprietary CRISPR Cas12b nuclease editing technology for a specified number of gene editing targets to research, develop and commercialize engineered cell therapy products that (i) are directed to certain antigen targets, with respect to our allogeneic T cell programs, or (ii) comprise certain human cell types, with respect to our stem cell-derived programs. We are permitted to use the CRISPR Cas12b system to modify or introduce, ex vivo, selected genetic sequences with respect to licensed products. The Beam Agreement excludes any rights to base editing using the CRISPR Cas12b system.
Pursuant to the Beam Agreement, we have the option, for a period of one year from the effective date of the Beam Agreement, to select additional antigen targets, with respect to our allogeneic T cell programs, or human cell types, with respect to our stem cell-derived programs, in each case, upon our payment of an option payment of $10 million per antigen target or cell type. In addition, we may, for a period of three years from the effective date of the Beam Agreement, (i) elect to replace an antigen target, with respect to our allogeneic T cell programs, or human cell type, with respect to our stem cell-derived programs (Replacement Right) previously selected by us, and (ii) select new gene editing targets, or replace gene editing targets previously selected by us, with respect to any licensed product (Gene Nomination Right). In each case, our rights with respect to its exercise of the option, Replacement Right or Gene Nomination Right are subject to certain limitations.
Pursuant to the Beam Agreement, we paid Beam an upfront payment of $50 million. Additionally, with respect to each licensed product, we will be obligated to pay to Beam up to $65 million in specified developmental and commercial milestone payments. We will also be obligated to pay to Beam an aggregate royalty, including any royalty owed by Beam to its licensor, on a licensed product-by-licensed product and country-by-country basis, in the low- to mid-single digits, subject to reduction in certain circumstances, on net sales of each licensed product until the latest of (i) the expiration of certain patents covering such licensed product in the applicable country, (ii) the date on which any applicable regulatory exclusivity, including orphan drug, new chemical entity, data or pediatric exclusivity, with respect to such licensed product expires in such country, or (iii) the 10th anniversary of the first commercial sale of such licensed product in such country.
Unless earlier terminated by either party, the Beam Agreement will expire on a licensed product-by-licensed product and country-by-country basis upon the expiration of our payment obligations with respect to each licensed product thereunder. We may terminate the Beam Agreement in its entirety or on an antigen target-by-antigen target basis (with respect to licensed product applicable to our allogeneic T cell programs), on a cell type-by-cell type basis (with respect to licensed product applicable to our stem cell-derived programs), or on a licensed product-by-licensed product basis, in each case, upon (i) 90 days’ advance written notice, if such notice is provided prior to the first commercial sale of a licensed product, or (ii) 180 days’ advance written notice, if such notice is provided after the first commercial sale of a licensed product. Either party may terminate the Beam Agreement with written notice for the other party’s material breach if such breaching party fails to timely cure the breach with respect to the country in which such material breach relates. Beam may terminate the Beam Agreement in its entirety if we or our affiliates or sublicensees commence a legal action challenging the validity, patentability, enforceability, or scope of any of the patent rights licensed to us thereunder. Either party also may terminate the Beam Agreement in its entirety upon certain insolvency events involving the other party.
License Agreement with the NIH
In January 2022, we entered into a patent license agreement (the NIH Agreement) with the U.S. Department of Health and Human Services, as represented by The National Cancer Institute, an institute of the National Institutes of Health (the NIH), pursuant to which the NIH granted to us an exclusive, worldwide, commercial license under certain patent rights related to certain fully-human anti-CD22 binders and CD22 CAR constructs comprising such binders for use in certain in vivo gene therapy and ex vivo allogeneic CAR T cell applications for B cell malignancies. The license grant is subject to customary statutory requirements and reserved rights as required under federal law and NIH requirements. We have the right to grant sublicenses under the licensed patent rights with the NIH’s prior consent.
Pursuant to the NIH Agreement, we paid to the NIH an upfront payment of $1.0 million. Additionally, we will be obligated to pay to the NIH (i) up to an aggregate of $9.6 million in specified regulatory, developmental, and commercial milestone payments
68
with respect to each product developed through exploitation of the licensed patent rights, and (ii) a payment of $1,000,000 upon the assignment of the NIH Agreement to an affiliate upon a change of control. In addition, we are obligated to pay to the NIH (i) a royalty on net sales of licensed products in the low-single digits, subject to reduction in certain circumstances, and subject to certain annual minimum royalty payments, and (ii) a percentage, ranging from the mid-single digits to mid-teens, of revenues from sublicensing arrangements. Additionally, if we are granted a priority review voucher by the U.S. Food and Drug Administration with respect to a licensed product, we will be obligated to pay to the NIH the greater of (i) $5,000,000 or (ii) a percentage in the mid-single digits of any consideration received for the sale, transfer, or lease of such priority review voucher. We are also obligated to pay to the NIH a percentage in the low-single digits of the consideration we receive for any assignment of the NIH Agreement to a non-affiliate.
We are obligated to use commercially reasonable efforts to exploit, and make publicly available, inventions developed by the exploitation of the licensed patent rights, including licensed products.
Unless earlier terminated by either party, the NIH Agreement will terminate upon expiration of the last-to-expire valid claim in the licensed patent rights. The NIH may terminate the Agreement with written notice for our material breach if we fail to timely cure such breach or upon certain insolvency events involving us. In addition, the NIH may terminate or modify the NIH Agreement, at its option, if the NIH determines that such termination or modification is necessary to meet the requirements for public use specified by federal regulations issued after the effective date of the NIH Agreement, and we do not reasonably and timely satisfy these requirements. We may terminate the NIH Agreement or any licenses in any country or territory upon 60 days’ prior written notice.
In Vivo Cell Engineering Platform
Cobalt Acquisition
In February 2019, we acquired all of the outstanding equity interests in Cobalt Biomedicine, Inc. (Cobalt), a privately-held early-stage biotechnology company founded by a Flagship Labs innovation team within Flagship Pioneering led by Dr. Geoffrey von Maltzahn that was developing a fusogen technology platform to specifically and consistently deliver diverse payloads—including DNA, RNA, and proteins—to targeted cells in vivo, in consideration of the issuance of 36.4 million shares of our Series A-2 convertible preferred stock, valued at $136.0 million. Of the 36.4 million shares of Series A-2 convertible preferred stock issued, 12.1 million shares were contingent on the achievement of a pre-specified development milestone, which was achieved in July 2019. We also agreed to pay contingent consideration of up to an aggregate of $500.0 million upon our achievement of certain pre-specified development milestones and a success payment of up to $500.0 million (the Cobalt Success Payment), which we may elect to pay in cash or in stock. The payout of the Cobalt Success Payment will only be paid if, at pre-determined valuation measurement dates, our market capitalization equals or exceeds $8.1 billion, and we are advancing a program based on the fusogen technology in a clinical trial pursuant to an IND, or have filed for, or received approval for, a BLA or NDA with respect to a program based on the fusogen technology. The valuation measurement dates for the Cobalt Success Payments are triggered by certain pre-determined valuation measurement dates, including the closing of our IPO and periodically thereafter. In addition to our IPO, a valuation measurement date would be triggered upon a change of control if at least one of our programs based on the fusogen technology is the subject of an active research program at the time of such change of control. As a result of the Cobalt transaction, we obtained licenses to various technologies and intellectual property rights that relate to the development of our fusogen technology and related fusosome programs, including exclusive license agreements with Flagship Pioneering Innovations V, Inc. (Flagship) and La Societe Pulsalys (Pulsalys), as well as several exclusive options to enter into exclusive license agreements, including one such option with The Regents of the University of California acting through The Technology Development Group of the University of California, Los Angeles (UCLA), with whom we later entered into an exclusive license agreement.
License Agreement with Flagship
In February 2016, Cobalt entered into an agreement (as amended, the Flagship Agreement), with Flagship, pursuant to which (i) Cobalt irrevocably and unconditionally assigned to Flagship all of its right, title and interest in and to certain foundational intellectual property developed by Flagship Pioneering, Inc. (Flagship Management) during the exploration and/or proto-company phase of Cobalt prior to its spin-out from Flagship (the Managerial Agreement), as set forth in the Flagship Agreement (such foundational intellectual property, the Fusogen Foundational IP), and (ii) Cobalt obtained an exclusive, worldwide, royalty-bearing, sublicensable, transferable license from Flagship under such Fusogen Foundational IP to develop, manufacture, and commercialize any product or process or component thereof, the development, manufacturing and commercialization of which would infringe at least one valid claim of Fusogen Foundational IP absent the license granted under the Flagship Agreement (Fusogen Products) in the field of human therapeutics during the term of the Flagship Agreement. In addition, Flagship irrevocably and unconditionally assigned to Cobalt all of its right, title and interest in and to any and all patents claiming any inventions conceived (i) solely by Flagship Management or jointly by Flagship Management and Cobalt, (ii) after Cobalt’s spinout from Flagship, and (iii) as a result of activities conducted pursuant the Managerial Agreement or other participation of Flagship Management in Cobalt’s affairs, but excluding Fusogen Foundational IP. We utilize the rights granted by Flagship under the Flagship Agreement in our fusogen platform and related
69
therapeutic product candidates. The license granted to Fusogen Foundational IP is contingent upon Cobalt’s compliance with its obligations under the Flagship Agreement. Under the Flagship Agreement, Cobalt also granted Flagship a non-exclusive, worldwide, royalty-free, fully paid, sublicensable license to practice the Fusogen Foundational IP within the field of human therapeutics solely to perform under the Managerial Agreement.
Pursuant to the Flagship Agreement, Cobalt is obligated to pay, on a Fusogen Product-by-Fusogen Product and jurisdiction-by-jurisdiction basis, royalties in the low single-digit percentage on net sales of Fusogen Products. The Flagship Agreement will terminate on the last-to-expire royalty term, which is determined on a Fusogen Product-by-Fusogen Product and jurisdiction-by-jurisdiction basis, and is the earlier of (i) the expiration of the last valid claim of any Fusogen Foundational IP covering such Fusogen Product or (ii) the date on which the last applicable additional milestone payment has been made in accordance with that certain merger agreement under which we acquired Cobalt, which we expect to be in 2039. Upon expiration of the royalty term with respect to a Fusogen Product in any jurisdiction and payment in full of all amounts owed under the Flagship Agreement for such Fusogen Product, the license granted to us will automatically convert into a non-exclusive, fully paid-up license for such Fusogen Product in such jurisdiction. We have the right to terminate the Flagship Agreement in its entirety for convenience upon 60 days of written notice. Either party may terminate the Flagship Agreement upon a material breach by the other party that is not cured within 30 days after receiving written notice. Also, Flagship may terminate the Flagship Agreement (i) upon 30 days’ written notice if we cease to carry on our business with respect to the rights granted in the Flagship Agreement, (ii) upon written notice if we experience an event of bankruptcy, or (iii) immediately upon written notice if we challenge the validity, patentability, or enforceability of any Fusogen Foundational IP or participate in any such challenge.
Sublicense Agreement with Pulsalys
In August 2018, Cobalt entered into an exclusive sublicense agreement (as amended, the Pulsalys Agreement), with Pulsalys, which Cobalt assigned to us in May 2020, and pursuant to which we obtained an exclusive, worldwide, sublicensable sublicense from Pulsalys of the exclusive license granted to Pulsalys by École normale supérieure de Lyon (ENS Lyon) on behalf of itself and Institut National de la Santé et de la Recherche Médicale (Inserm), Centre National de la Recherche Scientifique (CNRS) and Université Claude Bernard Lyon 1 (collectively, the Co-Owners) under certain patent rights relating to methods to selectively modulate the activity of distinct subtypes of immune cells using engineered virus-like particles. In addition, Pulsalys granted us the first right to negotiate an exclusive license to patent rights covering certain improvements to the licensed patent rights that are owned or held by Pulsalys. We utilize the rights granted under the Pulsalys Agreement in our in vivo fusogenic platform and related fusosome programs. Under the Pulsalys Agreement, we are obligated to use commercially reasonable efforts to develop and commercialize licensed products, which efforts we can demonstrate by the achievement of the following diligence milestones: (i) incurring a minimum annual spend of $1.0 million for each of the five years after the effective date of the Pulsalys Agreement, and (ii) submitting an IND within five years of the effective date of the Pulsalys Agreement. Under the Pulsalys Agreement, the Co-Owners will retain the right to practice the licensed patent rights for non-commercial research purposes, alone or in collaboration with third parties.
Pursuant to the Pulsalys Agreement, Cobalt paid Pulsalys an upfront fee of 18,000 EUR. We are required to pay an annual license maintenance fee of 18,000 EUR until the first commercial sale of a licensed product. We are also required to pay Pulsalys up to an aggregate of 575,000 EUR upon the achievement of certain clinical and regulatory milestones for each of the first three distinct licensed products. In addition, we are obligated to pay an annual royalty in the low single-digits on net sales of the licensed products, with the royalty rate being subject to reduction upon certain events. Lastly, we are obligated to pay percentage annual fees on certain sublicense income in the low single-digits .
The Pulsalys Agreement will terminate on a country-by-country and licensed product-by-licensed product basis upon the expiration of the last-to-expire valid claim within the licensed patent rights covering the making, using, sale, and import of such licensed product in such country or any patent term extension or supplementary protection certificate thereof covering the sale of such licensed product in such country, which we expect to occur in 2037. We also have the right to terminate the Pulsalys Agreement in its entirety upon notice if we determine, in our sole discretion, that continued pursuit of development of the licensed patent rights is not feasible or desirable in the context of (i) the resources available to us or due to external factors such as competition, market forces, or access or license to other reasonably useful intellectual property, or (ii) a change of direction of our business focus. Either party may terminate the Pulsalys Agreement upon a material breach by the other party that is not cured within 90 days after receiving written notice thereof. Pulsalys may terminate the Pulsalys Agreement (i) in full in the case of we undergo a cessation of business, dissolution or voluntary liquidation, or (ii) in full or in part (x) if we challenge the validity of the licensed patents, provided that such termination will be with respect to the claims within the licensed patents that are the subject of such challenge, or (y) if we fail to achieve the diligence milestones, and if the parties have not extended such milestones after good faith negotiations, and subject to our ability to cure such failure within 90 days after notice of the same.
70
License Agreement with UCLA
In March 2019, we entered into a license agreement (as amended, the UCLA Agreement) with UCLA, upon the exercise of an option originally granted by UCLA to Cobalt in April 2018. Under the UCLA Agreement, UCLA granted us an exclusive, sublicensable, transferable (subject to certain conditions) license in the licensed territory in the field of human therapeutics under certain patent rights relating to certain virus envelope pseudotyped lentiviruses and methods of their use to (i) research, make, have made, use, sell, offer for sale, have sold and import licensed products and (ii) practice licensed methods for the purposes of researching, manufacturing, and using licensed products, but not to perform services for a fee. The licensed territory under the UCLA Agreement is all countries of the world in which the licensed patent rights have or will be filed. UCLA agreed not to grant any rights under the licensed patents regarding licensed methods to third parties without first offering us an opportunity to remove the restrictions regarding the use of licensed methods to perform services for a fee. In addition, we agreed not to commercialize any licensed product that is not administered directly to a patient for therapeutic purposes without first negotiating with UCLA for possible development milestones, royalties, or other payments applicable to such licensed products. We utilize the rights granted under the UCLA Agreement in our in vivo fusogenic platform and related fusosome programs. We are obligated to use commercially reasonable and diligent efforts to (i) develop licensed products, (ii) market licensed products, and (ii) manufacture and sell licensed products in quantities sufficient to meet market demands. We are also required to satisfy certain development and commercial milestones with respect to at least one licensed product that is administered directly to a patient for therapeutic purposes.
The license granted pursuant to the UCLA Agreement is subject to certain rights retained by the California Institute for Regenerative Medicine (CIRM) and the U.S. government, including a non-exclusive, royalty-free license granted to the U.S. government in accordance with 35 U.S.C. §200-212. If CIRM exercises its rights under Title 17, California Code of Regulations, Section 100600, and the scope of our exclusive license under the UCLA Agreement is impacted, then our financial obligations therein will be reduced by 50%. Otherwise, rights retained by CIRM do not limit our ability to pursue our programs and product candidates. In addition, UCLA retains the right to (i) use the licensed patent rights for educational and research purposes and research sponsored by commercial entities, (ii) publicly disclose research results, (iii) use the licensed patent rights to offer and perform clinical diagnostic and prognostic care solely within the University of California system, and (iv) allow other non-profit and academic institutions to use the licensed patent rights for educational and research purposes and research sponsored by commercial entities, as well as to publicly disclose research results.
Pursuant to the UCLA Agreement, we paid UCLA an upfront license issue fee of $25,000. We also reimbursed UCLA for its past patent costs, and we have a continuing obligation to reimburse UCLA for its patent costs during the term of the UCLA Agreement. For licensed products that are administered directly to a patient for therapeutic purposes, we are required to pay UCLA up to an aggregate of (i) $825,000 upon the achievement of certain pre-specified development milestones for each of the first three such licensed products, and (ii) $15.0 million upon the achievement of certain pre-specified commercial milestones for such licensed products. In addition, we are obligated to pay an annual license maintenance fee beginning on the first anniversary of the UCLA Agreement until the first commercial sale of a licensed product. The license maintenance fee for the first anniversary was $10,000, and it will subsequently increase by $10,000 per anniversary up to a maximum annual license maintenance fee of $100,000. We are also required to pay, on a country-by-country basis, earned royalty percentages in the low single-digits on net sales of the licensed products, with the royalty rate being subject to reduction upon certain events. Under the UCLA Agreement, we are obligated to pay a minimum annual royalty of $100,000 beginning with the first full calendar year after the first commercial sale of a licensed product, and the minimum annual royalty will be credited against the earned royalty made during the same calendar year. If any claim within the licensed patent rights is held invalid or unenforceable in a final decision by a court of competent jurisdiction, all royalty obligations with respect to that claim or any claim patentably indistinct from it will expire as of the date of that final decision. No royalties will be collected or paid on licensed products sold to the U.S. government to the extent required by law, and we will be required to reduce the amount charged for licensed products distributed to the U.S. government by the amount of the royalty that otherwise would have been paid. Furthermore, we are obligated to pay UCLA tiered fees on a percentage of certain sublicense income in the low single-digit to low double-digit range. Lastly, if we challenge the validity of any licensed patent rights, we agree to pay UCLA all royalties and other amounts due in view of our activities under the UCLA Agreement during the period of challenge. If we fail such challenge, we are required to pay two times the royalty rate paid during the period of such challenge for the remaining term of the UCLA Agreement and all of UCLA’s verifiable legal out-of-pocket fees and costs incurred in defending such challenge, including attorney’s fees.
The UCLA Agreement will terminate on the later of the expiration of the last-to-expire patent or last to be abandoned patent application in the licensed patent rights, which we expect to occur in 2033. We also have the right to terminate the UCLA Agreement in its entirety or with respect to any portion of the licensed patent rights for any reason upon 90 days’ prior written notice to UCLA. UCLA may terminate the UCLA Agreement upon a material breach by us that is not cured within 90 days after receiving written notice. If the breach is incapable of being cured within such period, then UCLA will consider our efforts to avoid, and to take reasonable steps to cure, such breach when determining whether to terminate the UCLA Agreement. Also, UCLA has the right and option, at its sole discretion, to either terminate the UCLA Agreement or reduce our exclusive license to a non-exclusive license if we fail to (i) exercise commercially reasonable and diligent efforts to develop, market, manufacture, and sell licensed products, or
71
(ii) achieve certain development milestones set forth in the UCLA Agreement, subject to our ability to extend such milestones in accordance with terms set forth in the UCLA Agreement. Upon our termination of the UCLA Agreement, we may continue to sell any previously manufactured licensed products for 180 days after the effective date of termination. Upon termination of the UCLA Agreement by UCLA for our failure to reimburse UCLA for certain patent costs after the applicable cure period, we may continue to sell all previously made licensed products for 180 days after the effective date of the notice of termination; however, this right is not available if the UCLA Agreement is terminated for any other cause.
Government Regulation
The FDA and other regulatory authorities at federal, state, and local levels, as well as in foreign countries, extensively regulate, among other things, the research, development, testing, manufacture, quality control, import, export, safety, effectiveness, labeling, packaging, storage, distribution, record keeping, approval, advertising, promotion, marketing, post-approval monitoring, and post-approval reporting of biologics such as those we are developing. We, along with third-party contractors, will be required to navigate the various preclinical, clinical and commercial approval requirements of the governing regulatory agencies of the countries in which we wish to conduct studies or seek approval or licensure of our product candidates. The process of obtaining regulatory approvals and the subsequent compliance with applicable federal, state, local and foreign statutes and regulations require the expenditure of substantial time and financial resources.
U.S. Biologics Regulation
In the United States, biological products are subject to regulation under the Federal Food, Drug, and Cosmetic Act, the Public Health Service Act, and other federal, state, local and foreign statutes and regulations. The process required by the FDA before biologics may be marketed in the United States generally involves the following:
|
|
• |
completion of preclinical laboratory tests and animal studies performed in accordance with the FDA’s Good Laboratory Practice requirements (GLPs); |
|
|
• |
submission to the FDA of an Investigational new drug application (IND), which must become effective before clinical trials may begin; |
|
|
• |
approval by an institutional review board (IRB), or ethics committee at each clinical site before the trial is commenced; |
|
|
• |
performance of adequate and well-controlled human clinical trials to establish the safety, purity and potency of the proposed biologic product candidate for its intended purpose; |
|
|
• |
preparation of and submission to the FDA of a biologics license application (BLA), after completion of all pivotal clinical trials; |
|
|
• |
satisfactory completion of an FDA Advisory Committee review, if applicable; |
|
|
• |
a determination by the FDA within 60 days of its receipt of a BLA to file the application for review; |
|
|
• |
satisfactory completion of an FDA pre-approval inspection of the manufacturing facility or facilities at which the proposed product is produced to assess compliance with current Good Manufacturing Practices (cGMP), and to assure that the facilities, methods and controls are adequate to preserve the biological product’s continued safety, purity and potency and, if applicable, to assess compliance with the FDA’s current Good Tissue Practice (cGTP) requirements for the use of human cellular and tissue products, and of selected clinical investigation sites to assess compliance with Good Clinical Practices (GCPs); and |
|
|
• |
FDA review and approval of the BLA to permit commercial marketing of the product for particular indications for use in the United States. |
Prior to beginning the first clinical trial with a product candidate in the United States, we must submit an IND to the FDA. An IND is a request for authorization from the FDA to administer an investigational new drug to humans. The central focus of an IND submission is on the general investigational plan and the protocol(s) for clinical studies. The IND also includes results of animal and in vitro studies assessing the toxicology, pharmacokinetics, pharmacology, and pharmacodynamic characteristics of the product; chemistry, manufacturing, and controls information; and any available human data or literature to support the use of the investigational product. An IND must become effective before human clinical trials may begin. The IND automatically becomes effective 30 days after receipt by the FDA, unless the FDA, within the 30-day time period, raises safety concerns or questions about the proposed clinical trial. In such a case, the IND may be placed on clinical hold and the IND sponsor and the FDA must resolve any outstanding concerns or questions before the clinical trial can begin. Submission of an IND therefore may or may not result in FDA authorization to begin a clinical trial.
72
In addition to the IND submission process, under the National Institutes of Health (NIH), Guidelines for Research Involving Recombinant DNA Molecules, or the NIH Guidelines, supervision of human gene transfer trials includes evaluation and assessment by an institutional biosafety committee (IBC), a local institutional committee that reviews and oversees research utilizing recombinant or synthetic nucleic acid molecules at that institution. The IBC assesses the safety of the research and identifies any potential risk to public health or the environment, and such review may result in some delay before initiation of a clinical trial. While the NIH Guidelines are not mandatory unless the research in question is being conducted at or sponsored by institutions receiving NIH funding of recombinant or synthetic nucleic acid molecule research, many companies and other institutions not otherwise subject to the NIH Guidelines voluntarily follow them.
Clinical trials involve the administration of the investigational product to human subjects under the supervision of qualified investigators in accordance with GCPs, which include the requirement that all research subjects provide their informed consent for their participation in any clinical study. Clinical trials are conducted under protocols detailing, among other things, the objectives of the study, the parameters to be used in monitoring safety and the effectiveness criteria to be evaluated. A separate submission to the existing IND must be made for each successive clinical trial conducted during product development and for any subsequent protocol amendments. Furthermore, an independent IRB for each site proposing to conduct the clinical trial must review and approve the plan for any clinical trial and its informed consent form before the clinical trial begins at that site, and must monitor the study until completed. Regulatory authorities, the IRB or the sponsor may suspend a clinical trial at any time on various grounds, including a finding that the subjects are being exposed to an unacceptable health risk or that the trial is unlikely to meet its stated objectives. Some studies also include oversight by an independent group of qualified experts organized by the clinical study sponsor, known as a data safety monitoring board, which provides authorization for whether or not a study may move forward at designated check points based on access to certain data from the study and may halt the clinical trial if it determines that there is an unacceptable safety risk for subjects or other grounds, such as no demonstration of efficacy. There are also requirements governing the reporting of ongoing clinical studies and clinical study results to public registries.
For purposes of BLA approval, human clinical trials are typically conducted in three sequential phases that may overlap or be combined:
|
|
• |
Phase 1—The investigational product is initially introduced into healthy human subjects or patients with the target disease or condition. These studies are designed to test the safety, dosage tolerance, absorption, metabolism and distribution of the investigational product in humans, the side effects associated with increasing doses, and, if possible, to gain early evidence on effectiveness. |
|
|
• |
Phase 2—The investigational product is administered to a limited patient population with a specified disease or condition to evaluate the preliminary efficacy, optimal dosages and dosing schedule and to identify possible adverse side effects and safety risks. Multiple Phase 2 clinical trials may be conducted to obtain information prior to beginning larger and more expensive Phase 3 clinical trials. |
|
|
• |
Phase 3—The investigational product is administered to an expanded patient population to further evaluate dosage, to provide statistically significant evidence of clinical efficacy and to further test for safety, generally at multiple geographically dispersed clinical trial sites. These clinical trials are intended to establish the overall risk/benefit ratio of the investigational product and to provide an adequate basis for product approval. |
In some cases, the FDA may require, or companies may voluntarily pursue, additional clinical trials after a product is approved to gain more information about the product. These so-called Phase 4 studies may also be made a condition to approval of the BLA. Concurrent with clinical trials, companies may complete additional animal studies and develop additional information about the biological characteristics of the product candidate, and must finalize a process for manufacturing the product in commercial quantities in accordance with cGMP requirements. The manufacturing process must be capable of consistently producing quality batches of the product candidate and, among other things, must develop methods for testing the identity, strength, quality and purity of the final product. Additionally, appropriate packaging must be selected and tested, and stability studies must be conducted to demonstrate that the product candidate does not undergo unacceptable deterioration over its shelf life.
BLA Submission and Review by the FDA
Assuming successful completion of all required testing in accordance with all applicable regulatory requirements, the results of product development, nonclinical studies and clinical trials are submitted to the FDA as part of a BLA requesting approval to market the product for one or more indications. The BLA must include all relevant data available from preclinical and clinical studies, including negative or ambiguous results as well as positive findings, together with detailed information relating to the product’s chemistry, manufacturing, controls, and proposed labeling, among other things. Data can come from company-sponsored clinical studies intended to test the safety and effectiveness of a use of the product, or from a number of alternative sources, including studies initiated by independent investigators. The submission of a BLA requires payment of a substantial application user fee to the FDA, unless a waiver or exemption applies.
73
Within 60 days following submission of the application, the FDA reviews a BLA submitted to determine if it is substantially complete before the FDA accepts it for filing. The FDA may refuse to file any BLA that it deems incomplete or not properly reviewable at the time of submission and may request additional information. In this event, the BLA must be resubmitted with the additional information. Once a BLA has been accepted for filing, the FDA’s goal is to review standard applications within ten months after the filing date, or, if the application qualifies for priority review, six months after the FDA accepts the application for filing. In both standard and priority reviews, the review process may also be extended by FDA requests for additional information or clarification. The FDA reviews a BLA to determine, among other things, whether a product is safe, pure and potent and the facility in which it is manufactured, processed, packed or held meets standards designed to assure the product’s continued safety, purity and potency. The FDA may also convene an advisory committee to provide clinical insight on application review questions. The FDA is not bound by the recommendations of an advisory committee, but it considers such recommendations carefully when making decisions.
Before approving a BLA, the FDA will typically inspect the facility or facilities where the product is manufactured. The FDA will not approve an application unless it determines that the manufacturing processes and facilities are in compliance with cGMP and adequate to assure consistent production of the product within required specifications. For a product candidate that is also a human cellular or tissue product, the FDA also will not approve the application if the manufacturer is not in compliance with cGTPs. These are FDA regulations that govern the methods used in, and the facilities and controls used for, the manufacture of human cells, tissues, and cellular and tissue based products (HCT/Ps) which are human cells or tissue intended for implantation, transplant, infusion, or transfer into a human recipient. The primary intent of the GTP requirements is to ensure that cell and tissue based products are manufactured in a manner designed to prevent the introduction, transmission and spread of communicable disease. FDA regulations also require tissue establishments to register and list their HCT/Ps with the FDA and, when applicable, to evaluate donors through screening and testing. Additionally, before approving a BLA, the FDA will typically inspect one or more clinical sites to assure compliance with GCP. If the FDA determines that the application, manufacturing process or manufacturing facilities are not acceptable, it will outline the deficiencies in the submission and often will request additional testing or information. Notwithstanding the submission of any requested additional information, the FDA ultimately may decide that the application does not satisfy the regulatory criteria for approval.
After the FDA evaluates a BLA and conducts inspections of manufacturing facilities where the investigational product and/or its drug substance will be produced, the FDA may issue an approval letter or a Complete Response Letter (CRL). An approval letter authorizes commercial marketing of the product with specific prescribing information for specific indications. A CRL will describe all of the deficiencies that the FDA has identified in the BLA, except that where the FDA determines that the data supporting the application are inadequate to support approval, the FDA may issue the CRL without first conducting required inspections, testing submitted product lots, and/or reviewing proposed labeling. In issuing the CRL, the FDA may recommend actions that the applicant might take to place the BLA in condition for approval, including requests for additional information or clarification. The FDA may delay or refuse approval of a BLA if applicable regulatory criteria are not satisfied, require additional testing or information and/or require post-marketing testing and surveillance to monitor safety or efficacy of a product.
If regulatory approval of a product is granted, such approval will be granted for particular indications and may entail limitations on the indicated uses for which such product may be marketed. For example, the FDA may approve the BLA with a Risk Evaluation and Mitigation Strategy (REMS), to ensure the benefits of the product outweigh its risks. A REMS is a safety strategy implemented to manage a known or potential serious risk associated with a product and to enable patients to have continued access to such medicines by managing their safe use, and could include medication guides, physician communication plans, or elements to assure safe use, such as restricted distribution methods, patient registries and other risk minimization tools. The FDA also may condition approval on, among other things, changes to proposed labeling or the development of adequate controls and specifications. Once approved, the FDA may withdraw the product approval if compliance with pre- and post-marketing requirements is not maintained or if problems occur after the product reaches the marketplace. The FDA may require one or more Phase 4 post-market studies and surveillance to further assess and monitor the product’s safety and effectiveness after commercialization, and may limit further marketing of the product based on the results of these post-marketing studies.
Expedited development and review programs
The FDA offers a number of expedited development and review programs for qualifying product candidates. For example, the fast track program is intended to expedite or facilitate the process for reviewing new products that are intended to treat a serious or life-threatening disease or condition and demonstrate the potential to address unmet medical needs for the disease or condition. Fast track designation applies to the combination of the product and the specific indication for which it is being studied. The sponsor of a fast track product has opportunities for more frequent interactions with the applicable FDA review team during product development and, once a BLA is submitted, the product candidate may be eligible for priority review. A fast track product may also be eligible for rolling review, where the FDA may consider for review sections of the BLA on a rolling basis before the complete application is submitted, if the sponsor provides a schedule for the submission of the sections of the BLA, the FDA agrees to accept sections of the
74
BLA and determines that the schedule is acceptable, and the sponsor pays any required user fees upon submission of the first section of the BLA.
A product candidate intended to treat a serious or life-threatening disease or condition may also be eligible for breakthrough therapy designation to expedite its development and review. A product candidate can receive breakthrough therapy designation if preliminary clinical evidence indicates that the product candidate, alone or in combination with one or more other drugs or biologics, may demonstrate substantial improvement over existing therapies on one or more clinically significant endpoints, such as substantial treatment effects observed early in clinical development. The designation includes all of the fast track program features, as well as more intensive FDA interaction and guidance beginning as early as Phase 1 and an organizational commitment to expedite the development and review of the product candidate, including involvement of senior managers.
Any marketing application for a drug or biologic submitted to the FDA for approval, including a product candidate with a fast track designation and/or breakthrough therapy designation, may be eligible for other types of FDA programs intended to expedite the FDA review and approval process, such as priority review and accelerated approval. A product candidate is eligible for priority review if it is designed to treat a serious or life-threatening disease or condition, and if approved, would provide a significant improvement in safety or effectiveness compared to available alternatives for such disease or condition. For original BLAs, priority review designation means the FDA’s goal is to take action on the marketing application within six months of the 60-day filing date (as compared to ten months under standard review).
Additionally, product candidates studied for their safety and effectiveness in treating serious or life-threatening diseases or conditions may receive accelerated approval upon a determination that the product has an effect on a surrogate endpoint that is reasonably likely to predict clinical benefit, or on a clinical endpoint that can be measured earlier than irreversible morbidity or mortality, that is reasonably likely to predict an effect on irreversible morbidity or mortality or other clinical benefit, taking into account the severity, rarity, or prevalence of the condition and the availability or lack of alternative treatments. As a condition of accelerated approval, the FDA will generally require the sponsor to perform adequate and well-controlled post-marketing clinical studies to verify and describe the anticipated effect on irreversible morbidity or mortality or other clinical benefit. Products receiving accelerated approval may be subject to expedited withdrawal procedures if the sponsor fails to conduct the required post-marketing studies or if such studies fail to verify the predicted clinical benefit. In addition, the FDA currently requires as a condition for accelerated approval pre-approval of promotional materials, which could adversely impact the timing of the commercial launch of the product.
In 2017, the FDA established a new regenerative medicine advanced therapy (RMAT), designation as part of its implementation of the 21st Century Cures Act. The RMAT designation program is intended to fulfill the 21st Century Cures Act requirement that the FDA facilitate an efficient development program for, and expedite review of, any drug or biologic that meets the following criteria: (i) the drug or biologic qualifies as a RMAT, which is defined as a cell therapy, therapeutic tissue engineering product, human cell and tissue product, or any combination product using such therapies or products, with limited exceptions; (ii) the drug or biologic is intended to treat, modify, reverse, or cure a serious or life-threatening disease or condition; and (iii) preliminary clinical evidence indicates that the drug or biologic has the potential to address unmet medical needs for such a disease or condition. RMAT designation provides all the benefits of breakthrough therapy designation, including more frequent meetings with the FDA to discuss the development plan for the product candidate and eligibility for rolling review and priority review. Product candidates granted RMAT designation may also be eligible for accelerated approval on the basis of a surrogate or intermediate endpoint reasonably likely to predict long-term clinical benefit, or reliance upon data obtained from a meaningful number of clinical trial sites, including through expansion of trials to additional sites.
Fast track designation, breakthrough therapy designation, priority review, accelerated approval, and RMAT designation do not change the standards for approval but may expedite the development or approval process. Even if a product candidate qualifies for one or more of these programs, the FDA may later decide that the product no longer meets the conditions for qualification or decide that the time period for FDA review or approval will not be shortened.
Orphan drug designation and exclusivity
Under the Orphan Drug Act, the FDA may grant orphan designation to a drug or biologic intended to treat a rare disease or condition, defined as a disease or condition with a patient population of fewer than 200,000 individuals in the United States, or a patient population greater than 200,000 individuals in the United States and when there is no reasonable expectation that the cost of developing and making available the drug or biologic in the United States will be recovered from sales in the United States for that drug or biologic. Orphan drug designation must be requested before submitting a BLA. After the FDA grants orphan drug designation, the generic identity of the therapeutic agent and its potential orphan use are disclosed publicly by the FDA.
If a product that has orphan drug designation subsequently receives the first FDA approval for a particular active ingredient for the disease for which it has such designation, the product is entitled to orphan product exclusivity, which means that the FDA may not
75
approve any other applications, including a full BLA, to market the same biologic for the same indication for seven years, except in limited circumstances, such as a showing of clinical superiority to the product with orphan drug exclusivity or if the FDA finds that the holder of the orphan drug exclusivity has not shown that it can assure the availability of sufficient quantities of the orphan drug to meet the needs of patients with the disease or condition for which the drug was designated. Orphan drug exclusivity does not prevent the FDA from approving a different drug or biologic for the same disease or condition, or the same drug or biologic for a different disease or condition. Among the other benefits of orphan drug designation are tax credits for certain research and a waiver of the BLA application user fee.
A designated orphan drug may not receive orphan drug exclusivity if it is approved for a use that is broader than the indication for which it received orphan designation. In addition, orphan drug exclusive marketing rights in the United States may be lost if the FDA later determines that the request for designation was materially defective or, as noted above, if a second applicant demonstrates that its product is clinically superior to the approved product with orphan exclusivity or the manufacturer of the approved product is unable to assure sufficient quantities of the product to meet the needs of patients with the rare disease or condition.
Post-approval requirements
Biologics are subject to pervasive and continuing regulation by the FDA, including, among other things, requirements relating to record-keeping, reporting of adverse experiences, periodic reporting, product sampling and distribution, and advertising and promotion of the product. After approval, most changes to the approved product, such as adding new indications or other labeling claims, are subject to prior FDA review and approval. There also are continuing, annual program fees for any marketed products. Biologic manufacturers and their subcontractors are required to register their establishments with the FDA and certain state agencies, and are subject to periodic unannounced inspections by the FDA and certain state agencies for compliance with cGMP, which impose certain procedural and documentation requirements up. Changes to the manufacturing process are strictly regulated, and, depending on the significance of the change, may require prior FDA approval before being implemented. FDA regulations also require investigation and correction of any deviations from cGMP and impose reporting requirements. Accordingly, manufacturers must continue to expend time, money and effort in the area of production and quality control to maintain compliance with cGMP and other aspects of regulatory compliance.
The FDA may withdraw approval if compliance with regulatory requirements and standards is not maintained or if problems occur after the product reaches the market. Later discovery of previously unknown problems with a product, including adverse events of unanticipated severity or frequency, or with manufacturing processes, or failure to comply with regulatory requirements, may result in revisions to the approved labeling to add new safety information; imposition of post-market studies or clinical studies to assess new safety risks; or imposition of distribution restrictions or other restrictions under a REMS program. Other potential consequences include, among other things:
|
|
• |
restrictions on the marketing or manufacturing of the product, complete withdrawal of the product from the market or product recalls; |
|
|
• |
fines, warning letters, or untitled letters; |
|
|
• |
clinical holds on clinical studies; |
|
|
• |
refusal of the FDA to approve pending applications or supplements to approved applications, or suspension or revocation of product license approvals; |
|
|
• |
product seizure or detention, or refusal to permit the import or export of products; |
|
|
• |
consent decrees, corporate integrity agreements, debarment or exclusion from federal healthcare programs; |
|
|
• |
mandated modification of promotional materials and labeling and the issuance of corrective information; |
|
|
• |
the issuance of safety alerts, Dear Healthcare Provider letters, press releases and other communications containing warnings or other safety information about the product; or |
|
|
• |
injunctions or the imposition of civil or criminal penalties. |
The FDA closely regulates the marketing, labeling, advertising and promotion of biologics. A company can make only those claims relating to safety and efficacy, purity and potency that are approved by the FDA and in accordance with the provisions of the approved label. The FDA and other agencies actively enforce the laws and regulations prohibiting the promotion of off-label uses. Failure to comply with these requirements can result in, among other things, adverse publicity, warning letters, corrective advertising and potential civil and criminal penalties. Physicians may prescribe legally available products for uses that are not described in the product’s labeling and that differ from those tested and approved by the FDA. Such off-label uses are common across medical specialties. Physicians may believe that such off-label uses are the best treatment for many patients in varied circumstances. The FDA
76
does not regulate the behavior of physicians in their choice of treatments. The FDA does, however, restrict manufacturer’s communications on the subject of off-label use of their products.
Biosimilars and reference product exclusivity
The Affordable Care Act, signed into law in 2010, includes a subtitle called the Biologics Price Competition and Innovation Act (BPCIA), which created an abbreviated approval pathway for biological products that are biosimilar to or interchangeable with an FDA-licensed reference biological product. The FDA has issued several guidance documents outlining an approach to review and approval of biosimilars.
Biosimilarity, which requires that there be no clinically meaningful differences between the biological product and the reference product in terms of safety, purity, and potency, can be shown through analytical studies, animal studies, and a clinical study or studies. Interchangeability requires that a product is biosimilar to the reference product and the product must demonstrate that it can be expected to produce the same clinical results as the reference product in any given patient and, for products that are administered multiple times to an individual, the biologic and the reference biologic may be alternated or switched after one has been previously administered without increasing safety risks or risks of diminished efficacy relative to exclusive use of the reference biologic. However, complexities associated with the larger, and often more complex, structures of biological products, as well as the processes by which such products are manufactured, pose significant hurdles to implementation of the abbreviated approval pathway that are still being worked out by the FDA.
Under the BPCIA, an application for a biosimilar product may not be submitted to the FDA until four years following the date that the reference product was first licensed by the FDA. In addition, the approval of a biosimilar product may not be made effective by the FDA until 12 years from the date on which the reference product was first licensed. During this 12-year period of exclusivity, another company may still market a competing version of the reference product if the FDA approves a full BLA for the competing product containing that applicant’s own preclinical data and data from adequate and well-controlled clinical trials to demonstrate the safety, purity and potency of its product. The BPCIA also created certain exclusivity periods for biosimilars approved as interchangeable products. At this juncture, it is unclear whether products deemed “interchangeable” by the FDA will, in fact, be readily substituted by pharmacies, which are governed by state pharmacy law.
A biological product can also obtain pediatric market exclusivity in the United States. Pediatric exclusivity, if granted, adds six months to existing exclusivity periods and patent terms. This six-month exclusivity, which runs from the end of other exclusivity protection or patent term, may be granted based on the voluntary completion of a pediatric study in accordance with an FDA-issued “Written Request” for such a study. The BPCIA is complex and continues to be interpreted and implemented by the FDA. In addition, government proposals have sought to reduce the 12-year reference product exclusivity period. Other aspects of the BPCIA, some of which may impact the BPCIA exclusivity provisions, have also been the subject of recent litigation. As a result, the ultimate impact, implementation, and impact of the BPCIA is subject to significant uncertainty.
Other Healthcare Laws
Pharmaceutical companies are subject to additional healthcare regulation and enforcement by the federal government and by authorities in the states and foreign jurisdictions in which they conduct their business and may constrain the financial arrangements and relationships through which we research, as well as, sell, market and distribute any products for which we obtain marketing approval. Such laws include, without limitation, federal and state anti-kickback, fraud and abuse, false claims, data privacy and security and physician and other health care provider transparency laws and regulations. If our significant operations are found to be in violation of any of such laws or any other governmental regulations that apply, they may be subject to penalties, including, without limitation, administrative, civil and criminal penalties, damages, fines, disgorgement, the curtailment or restructuring of operations, integrity oversight and reporting obligations, exclusion from participation in federal and state healthcare programs and imprisonment.
Coverage and Reimbursement
Sales of any product depend, in part, on the extent to which such product will be covered by third-party payors, such as federal, state, and foreign government healthcare programs, commercial insurance and managed healthcare organizations, and the level of reimbursement for such product by third-party payors. Decisions regarding the extent of coverage and amount of reimbursement to be provided are made on a plan-by-plan basis. These third-party payors are increasingly reducing reimbursements for medical products, drugs and services. In addition, the U.S. government, state legislatures and foreign governments have continued implementing cost-containment programs, including price controls, restrictions on coverage and reimbursement and requirements for substitution of generic products. Adoption of price controls and cost-containment measures, and adoption of more restrictive policies in jurisdictions with existing controls and measures, could further limit sales of any product. Decreases in third-party reimbursement for any product or a decision by a third-party payor not to cover a product could reduce physician usage and patient demand for the product and also have a material adverse effect on sales.
77
Healthcare Reform
In March 2010, the Patient Protection and Affordable Care Act, as amended by the Health Care and Education Reconciliation Act, each as amended, collectively known as the ACA, was enacted, which substantially changed the way healthcare is financed by both governmental and private insurers, and significantly affected the pharmaceutical industry. The ACA contains a number of provisions, including those governing enrollment in federal healthcare programs, reimbursement adjustments and changes to fraud and abuse laws. For example, the ACA:
|
|
• |
increased the minimum level of Medicaid rebates payable by manufacturers of brand name drugs from 15.1% to 23.1% of the average manufacturer price; |
|
|
• |
required collection of rebates for drugs paid by Medicaid managed care organizations; |
|
|
• |
required manufacturers to participate in a coverage gap discount program, under which they must agree to offer 70 percent point-of-sale discounts off negotiated prices of applicable brand drugs to eligible beneficiaries during their coverage gap period, as a condition for the manufacturer’s outpatient drugs to be covered under Medicare Part D; and |
|
|
• |
imposed a non-deductible annual fee on pharmaceutical manufacturers or importers who sell “branded prescription drugs” to specified federal government programs. |
Since its enactment, there have been judicial and Congressional challenges to certain aspects of the ACA, and we expect there will be additional challenges and amendments to the ACA in the future. For example, on March 2, 2020 the United States Supreme Court granted the petitions for writs of certiorari to review the U.S. Court of Appeals for the 5th Circuit ruling that the individual mandate was unconstitutional and to determine the constitutionality of the ACA in its entirety. It is uncertain when the Supreme Court will rule on this case. Other legislative changes have been proposed and adopted since the ACA was enacted, including aggregate reductions of Medicare payments to providers of 2% per fiscal year, which was temporarily suspended from May 1, 2020 through March 31, 2021 due to the COVID-19 pandemic, and reduced payments to several types of Medicare providers. Moreover, there has recently been heightened governmental scrutiny over the manner in which manufacturers set prices for their marketed products, which has resulted in several Congressional inquiries, proposed and enacted legislation and executive orders issued by the President designed to, among other things, bring more transparency to product pricing, review the relationship between pricing and manufacturer patient programs, and reform government program reimbursement methodologies for drug products. It is also possible that additional governmental action is taken in response to the COVID-19 pandemic. Individual states in the United States have also become increasingly active in implementing regulations designed to control pharmaceutical product pricing, including price or patient reimbursement constraints, discounts, restrictions on certain product access and marketing cost disclosure and transparency measures, and, in some cases, designed to encourage importation from other countries and bulk purchasing.
Employees and Human Capital Resources
As of December 31, 2021, we had 383 employees, 302 of whom were primarily engaged in research and development activities. A total of 236 employees have an advanced degree. None of our employees are represented by a labor union or party to a collective bargaining agreement. We consider our relationship with our employees to be good.
Our human capital resources objectives include, as applicable, identifying, recruiting, retaining, incentivizing and integrating our existing and additional employees. The principal purposes of our equity incentive plans are to attract, retain and motivate selected employees, consultants and directors through the granting of stock-based compensation awards and cash-based performance bonus awards.
Legal Proceedings
We are not currently a party to any material legal proceedings. From time to time, we may, however, in the ordinary course of business face various claims brought by third parties, and we may, from time to time, make claims or take legal actions to assert our rights, including intellectual property rights as well as claims relating to employment matters and the safety or efficacy of our products. Any of these claims could subject us to costly litigation, and, while we generally believe that we have adequate insurance to cover many different types of liabilities, our insurance carriers may deny coverage, may be inadequately capitalized to pay on valid claims, or our policy limits may be inadequate to fully satisfy any damage awards or settlements. If this were to happen, the payment of any such awards could have a material adverse effect on our operations, cash flows and financial position. Additionally, any such claims, whether or not successful, could damage our reputation and business.
78
Our Corporate Information
We were founded in July 2018 as a Delaware corporation. Our principal executive offices are located at 188 East Blaine Street, Suite 400, Seattle, Washington 98102, and our telephone number is (206) 701-7914. Our website address is www.sana.com. The information on, or that can be accessed through, our website is not part of this report, and is not incorporated by reference herein. We have included our website address as an inactive textual reference only. We may use our website as a means of disclosing material non-public information and for complying with our disclosure obligations under Regulation Fair Disclosure promulgated by the SEC. These disclosures will be included on our website under the “Investors” section.
79
Item 1A. Risk Factors.
Investing in shares of our common stock involves a high degree of risk. You should carefully consider the following risks and uncertainties, together with all of the other information contained in this Annual Report, including our financial statements and related notes included elsewhere in this Annual Report, before making an investment decision. The risks described below are not the only ones facing us. Moreover, we may have already experienced the circumstances described in one or more of the risk factors described below. Many of the following risks and uncertainties are, and will continue to be, exacerbated by the ongoing COVID-19 pandemic and any worsening of the global business and economic environment as a result. The occurrence of any of the following risks, or of additional risks and uncertainties not presently known to us or that we currently believe to be immaterial, could materially and adversely affect our business, financial condition, reputation, or results of operations. In such a case, the trading price of shares of our common stock could decline, and you may lose all or part of your investment.
Risks Related to Our Business and Industry
Our ex vivo and in vivo cell engineering platforms are based on novel technologies that are unproven and may not result in approvable or marketable products. This uncertainty exposes us to unforeseen risks, makes it difficult for us to predict the time and cost that will be required for the development and potential regulatory approval of our product candidates and increases the risk that we may ultimately not be successful in our efforts to use and expand our technology platforms to build a pipeline of product candidates.
We are seeking to identify and develop a broad pipeline of product candidates using our ex vivo and in vivo cell engineering platforms. We have not commenced clinical trials for any product candidates developed with these platforms. The scientific research that forms the basis of our efforts to develop product candidates with our platforms is still ongoing. We are not aware of any United Stated Food and Drug Administration (FDA)-approved therapeutics that utilize fusogen technology or that are cell products derived from pluripotent stem cells (PSCs). Further, the scientific evidence that supports the feasibility of developing therapeutic treatments based on our platforms is both preliminary and limited. As a result, we are exposed to a number of unforeseen risks, and it is difficult to predict the types of challenges and risks that we may encounter during development of our product candidates. For example, we have not tested our cell engineering platforms on all pluripotent and differentiated cell types or in all microenvironments, so results from one cell type or microenvironment may not translate into other cell types or microenvironments. In addition, our current gene editing approaches rely on novel gene editing reagents that may have unanticipated or undesirable effects or prove to be less effective than we expect. Also, we have not tested any of the product candidates that we are developing using our cell engineering platforms in humans, and our current data is limited to animal models and preclinical cell lines, the results of which may not translate into humans. Further, relevant animal models and assays may not accurately predict the safety and efficacy of our product candidates in humans, and we may encounter significant challenges creating appropriate models and assays for demonstrating the safety and purity of our product candidates.
In addition, our fusogen and hypoimmune technologies have potential safety risks, including those related to genotoxicity associated with the delivery of genome-modifying payloads. For example, DNA sequences that randomly integrate into a cell’s DNA may increase risk for or cause certain cancers. Alternatively, gene-editing approaches may edit the genome at sites other than the intended DNA target or cause DNA rearrangements, each of which may have oncogenic or other adverse effects. PSC-derived cell products may have potential safety risks related to genomic variations that have been observed during passage (i.e., amplification) and differentiation of pluripotent cell lines. We cannot always predict the types and potential impact of these genomic changes, including whether certain changes are harmful. Accordingly, it may be difficult for us to conduct the level of testing and development of assays necessary to ensure the safety of our PSC-derived cell product candidates in humans. These risks related to genetic variation are also relevant to our product candidates created from donor-derived cells. Additionally, our stem cell-based product candidates have potential safety risks related to insufficient cell differentiation that may lead to oncogenic transformations or other adverse effects. As a result, it is possible that safety events or concerns could negatively affect the development of our product candidates, including by adversely affecting patient enrollment in future clinical trials of our product candidates among the patient populations that we intend to treat.
Given the novelty of our technologies, we intend to work closely with the FDA and comparable foreign regulatory authorities to perform the requisite scientific analyses and evaluation of our methods to obtain regulatory approval for our product candidates. However, due to a lack of experience with similar therapeutics, the regulatory pathway with the FDA and comparable regulatory authorities may be more complex, time-consuming, and unpredictable relative to more well-known therapeutics. Even if we obtain human data to support continued evaluation and approval of our product candidates, the FDA or comparable foreign regulatory authorities may lack experience in evaluating the safety and efficacy of therapeutics similar to our product candidates. For example, given that there are no approved PSC- or donor-derived cell products on the market, the FDA and comparable foreign regulatory authorities have not established consistent standards by which to evaluate the safety of such products, and any such standards that they do establish may subsequently change. Moreover, the FDA has increased its focus in recent years on potential safety issues associated with gene and cell therapy products, including by placing clinical holds on certain product candidates pending further evaluation of genomic abnormalities detected in as few as a single patient following administration of such product candidates. We cannot be
80
certain that the FDA or comparable foreign regulatory authorities will determine that the potential safety risks associated with our PSC- or donor-derived cell product candidates outweigh the potential therapeutic benefits, and that they will allow us to commence clinical trials of such product candidates in a timely manner, or at all, or to continue such clinical trials once they have commenced. If we become subject to a clinical hold with respect to any of our product candidates due to a potential safety issue, we cannot guarantee that we will be able to provide the applicable regulatory authority with sufficient data or other evidence regarding the safety of such product candidate such that we can resume clinical development of such product candidates in a timely manner or at all. This could delay clinical development of such product candidate or our other product candidates, increase our expected development costs, increase the length of the regulatory review process, and delay or prevent commercialization of our product candidates. Moreover, even if we and the applicable regulatory authorities determine that our product candidates are safe in humans, and such products obtain approval, they may later prove to cause serious adverse side effects in patients that we were unable to observe or predict during the clinical development of such product candidates, which may subject us to significant negative consequences, as described elsewhere in these Risk Factors. In addition, the evaluation process for our product candidates takes time and resources and may require independent third-party analyses, and our product candidates may not be accepted or approved by the FDA or comparable foreign regulatory authorities. We cannot be certain that our ex vivo and in vivo cell engineering platforms will lead to the development of approvable or marketable products, either alone or in combination with other therapies.
Additionally, a key element of our strategy is to use and expand our ex vivo and in vivo cell engineering platforms to build a pipeline of product candidates and advance those product candidates through clinical development for the treatment of a variety of different types of diseases. Although our research and development efforts to date have been focused on identifying a pipeline of product candidates directed at various disease types, we may not be able to develop product candidates that are safe and effective. Even if we are successful in building our pipeline, the potential product candidates that we identify may not be suitable for clinical development, including if they are shown to have harmful side effects or other characteristics that indicate that they are unlikely to receive marketing approval and achieve market acceptance. If we do not successfully develop, obtain approval for, and commercialize any of our current or future product candidates, we will face difficulty in generating product revenue in future periods, which could result in significant harm to our financial position and adversely affect our share price.
If we are unable to successfully identify, develop, and commercialize any product candidates, or experience significant delays in doing so, our business, financial condition, and results of operations will be materially adversely affected.
Our ability to generate revenue from sales of any of our product candidates, which we do not expect to occur for at least the next several years, if ever, will depend heavily on the timely and successful identification, development, regulatory approval, and eventual commercialization of any such product candidates, which may never occur. To date, we have not generated revenue from sales of any products, and we may never be able to develop, obtain regulatory approval for, or commercialize a marketable product. All of our current product candidates are in preclinical development, and, before we generate any revenue from product sales, will require that we manage preclinical, clinical, and manufacturing activities, undertake significant clinical development, obtain regulatory approval in multiple jurisdictions, establish manufacturing supply, including commercial manufacturing supply, and build a commercial organization, which will require a substantial investment and significant marketing efforts. We may never receive regulatory approval for any of our product candidates, which would prevent us from marketing or promoting any of our product candidates.
The successful development of our product candidates will depend on numerous factors, including the following:
|
|
• |
our successful and timely completion of preclinical studies and clinical trials for which the FDA, and any comparable foreign regulatory authorities, agree with the design, endpoints, and implementation; |
|
|
• |
the sufficiency of our financial and other resources to complete the necessary preclinical studies and clinical trials; |
|
|
• |
our receipt of regulatory approvals or authorizations for conducting future clinical trials; |
|
|
• |
our ability to timely and successfully initiate, enroll patients in, and complete clinical trials; |
|
|
• |
our ability to demonstrate to the satisfaction of the FDA or any comparable foreign regulatory authority that the applicable product candidate is safe and efficacious, has suitable purity, and is potent as a treatment for our targeted indications; |
|
|
• |
our ability to demonstrate to the satisfaction of the FDA or any comparable foreign regulatory authority that the applicable product candidate’s risk-benefit ratio for its proposed indication is acceptable; |
|
|
• |
the timely receipt of marketing approvals for our product candidates from applicable regulatory authorities; |
|
|
• |
our ability to address any potential interruptions or delays resulting from factors related to the ongoing COVID-19 pandemic; |
|
|
• |
the extent of any required post-marketing approval commitments to applicable regulatory authorities, including the conduct of any post-marketing approval clinical studies, and our ability to comply with any such commitments; and |
81
|
|
• |
our ability to establish, scale up, and scale out, either alone or with third-party manufacturers, manufacturing capabilities for clinical supply of our product candidates for our clinical trials and, if any of our product candidates are approved, commercial supply (including licensure) of such product candidates. |
Additionally, clinical or regulatory setbacks experienced by other companies developing similar products or within adjacent fields, including allogeneic cell-based therapies and the fields of gene editing and gene therapy, may impact the clinical development of and regulatory pathway for our current or future product candidates or negatively impact the perceptions of value or risk of our technologies.
If we experience issues with or delays with respect to any one or more of these factors, we could experience significant delays or be unable to successfully develop and commercialize our product candidates, which would materially adversely affect our business, financial condition, and results of operations.
While we believe our pipeline will yield multiple INDs, we may not be able to submit INDs to commence clinical trials on the timelines we expect, and even if we are able to submit an IND, the FDA may not permit us to proceed with clinical trials.
We expect our pipeline to yield multiple Investigational New Drug applications (INDs) beginning as early as 2022, including INDs for our allogeneic CAR T cell product candidates from our ex vivo cell engineering platform and our fusosome CAR T product candidates from our in vivo cell engineering platform. We cannot be sure that, following our submission of an IND, the FDA or comparable foreign regulatory authorities will allow our clinical trials to begin, or that, once begun, issues will not arise that require suspension or termination of such clinical trials. The manufacturing of our product candidates, including our CAR T ex vivo cell engineering product candidates, remains an emerging and evolving field. Accordingly, we expect topics relating to chemistry, manufacturing, and controls, including product specifications, will be a focus of IND reviews, which may delay the clearance of INDs that we submit. Additionally, even if applicable regulatory authorities agree with the design and implementation of the clinical trials set forth in an IND or comparable foreign submission, such regulatory authorities may change their requirements in the future, which could require us to make costly changes to and delay the conduct of our clinical trials or require suspension or termination of such trials entirely.
We may not realize the benefits of technologies that we have acquired, or will acquire in the future, or other strategic transactions that we have or will consummate.
Our ex vivo and in vivo cell engineering technology represents an aggregation of years of innovation and technology from multiple academic institutions and companies, including our fusogen technology that we acquired from Cobalt, our ex vivo cell engineering programs focused on replacing damaged cells in the heart and certain brain disorders that we acquired from Cytocardia Inc. (Cytocardia) and Oscine Corp. (Oscine), respectively, hypoimmune technology that we licensed from Harvard and The Regents of the University of California (UCSF), and gene editing technology that we licensed from Beam Therapeutics Inc., among others. Further, a key component of our strategy is to acquire and in-license technologies to support our mission of using engineered cells as medicines. As such, we actively evaluate various strategic transactions on an ongoing basis. We may acquire other businesses, products, or technologies, as well as pursue joint ventures or investments in complementary businesses. The level of success of these strategic transactions, including any future strategic transactions, will depend on the risks and uncertainties involved, including:
|
|
• |
unanticipated liabilities related to acquired companies or joint ventures; |
|
|
• |
difficulties integrating acquired personnel, technologies, and operations into our existing business; |
|
|
• |
difficulty retaining key employees, including of any acquired businesses; |
|
|
• |
diversion of management time and focus from operating our business to management of acquisition and integration efforts, strategic alliances or collaborations, or joint venture challenges; |
|
|
• |
increases in our expenses and reductions in our cash available for operations and other uses; |
|
|
• |
higher than expected collaboration, acquisition, or integration costs; |
|
|
• |
disruption in our relationships with collaborators, key suppliers, manufacturers, or customers as a result of such transactions; |
|
|
• |
incurrence of substantial debt or dilutive issuances of equity securities to pay transaction consideration or costs; |
|
|
• |
possible write-offs of assets, goodwill or impairment charges, or increased amortization expenses relating to acquired businesses or joint ventures; |
|
|
• |
difficulty and cost in facilitating the collaboration or combining the operations and personnel of any acquired business; and |
82
|
|
• |
challenges resulting from the COVD-19 pandemic that make it more difficult to integrate acquired businesses into our business. |
In addition, foreign acquisitions and joint ventures are subject to additional risks, including those related to integration of operations across different cultures and languages, currency risks, potentially adverse tax consequences of overseas operations, and the particular economic, political, and regulatory risks associated with specific countries. The occurrence of any of these risks or uncertainties may preclude us from realizing the anticipated benefit of any acquisition or strategic transaction, and our financial condition may be harmed.
Additionally, we may not be successful in our efforts to acquire or obtain rights to certain technologies or products that are necessary for the success of our product candidates on acceptable terms or at all, including because we may be unable to successfully or timely negotiate the terms of an agreement with the third-party owner of such technology or products or because such third party may have determined to deprioritize such technology or products. If we are not able to acquire or obtain rights to certain technologies or products on which certain of our product candidates may depend, it may be necessary for us to curtail, reduce, or delay the development of such product candidates.
We may not realize the benefits of any collaborative or licensing arrangement, and if we fail to enter into new strategic relationships, our business, financial condition, commercialization prospects, and results of operations may be materially adversely affected.
Our product development programs and the potential commercialization of our product candidates will require substantial additional cash to fund expenses. In addition, our ex vivo and in vivo cell engineering platforms are attractive technologies for potential collaborations due to their breadth of application. Therefore, for certain of our product candidates, including product candidates that we may develop in the future, we may decide to form or seek strategic alliances, collaborations, or licensing arrangements with pharmaceutical or biotechnology companies that we believe will complement or augment our development and potential commercialization efforts with respect to such product candidates, including in territories outside the United States or for certain indications.
We face significant competition in seeking appropriate collaborators. Collaborations are complex and time-consuming to negotiate and document. We may not be successful in our efforts to establish a strategic partnership or other alternative arrangements for our product candidates on acceptable terms or at all, including because our product candidates may be deemed to be at too early of a stage of development for collaborative effort or third parties may not view our product candidates as having the requisite potential to demonstrate safety and efficacy. Additionally, there have been a significant number of recent business combinations among large pharmaceutical companies that have reduced the number of potential future collaborators and changed the strategies of the resulting combined companies. In addition, under the terms of certain license agreements applicable to our product candidates, we may be restricted from entering into agreements on certain terms or at all with potential collaborators relating to those product candidates. If and when we collaborate with a third party for development and commercialization of a product candidate, we expect that we may have to relinquish some or all of the control over the future success of that product candidate to the third party. Our ability to reach a definitive agreement for a collaboration will depend, among other things, upon our assessment of the collaborator’s resources and expertise, the terms and conditions of the proposed collaboration, and the proposed collaborator’s evaluation of our technologies, product candidates, and market opportunities. The collaborator may also consider alternative product candidates or technologies for similar indications that may be available for collaboration and could determine that such other collaboration is more attractive than a collaboration with us for our product candidate.
In instances where we do enter into collaborations, we could be subject to the following risks, each of which may materially harm our business, commercialization prospects, and financial condition:
|
|
• |
collaborators may have significant discretion in determining the efforts and resources that they will apply to a collaboration and may not commit sufficient efforts and resources to the product development or marketing programs or may misapply those efforts and resources; |
|
|
• |
collaborators may experience financial difficulties; |
|
|
• |
collaborators may not pursue development and commercialization of collaboration product candidates or may elect not to continue or renew development or commercialization programs based on clinical trial results or changes in their strategic focus; |
83
|
|
• |
collaborators may delay clinical trials, fail to provide sufficient funding for a clinical trial program, stop a clinical trial or abandon a product candidate, repeat or conduct new clinical trials, or require a new formulation of a product candidate for clinical testing; |
|
|
• |
we may be required to relinquish important rights to our product candidates, such as marketing, distribution, and intellectual property rights; |
|
|
• |
we may be required to agree to exclusivity, non-competition, or other terms that restrict our ability to research, develop, or commercialize certain existing product candidates or potential future product candidates, including our ability to develop our product candidates in certain indications or geographic regions or combine our product candidates with certain third-party products; |
|
|
• |
collaborators may not properly maintain or defend our intellectual property rights or may use our proprietary information in way that gives rise to actual or threatened litigation that could jeopardize or invalidate our intellectual property rights or proprietary information or expose us to litigation or potential liability; |
|
|
• |
collaborators may infringe the intellectual property rights of third parties, which may expose us to litigation and potential liability; |
|
|
• |
collaborators may acquire outside of the collaboration or develop, independently or in collaboration with third parties, including our competitors, products that compete directly or indirectly with our products or product candidates and may move forward with such products instead of ours; |
|
|
• |
collaborators may own or co-own intellectual property rights covering our products that result from our collaboration, and in such cases, we may not have an exclusive right to commercialize the product candidates covered by such intellectual property rights. |
|
|
• |
we and our collaborators may disagree regarding the development plan for a product candidate with respect to which we are collaborating, including, for example, with respect to target indications, inclusion or exclusion criteria for a clinical trial, or the decision to seek front line therapy approval versus second-, third-, or fourth-line therapy approval; |
|
|
• |
disputes may arise between the collaborators and us that result in the delay or termination of the research, development, or commercialization of our product candidates or that may result in costly litigation or arbitration that diverts management attention and resources; |
|
|
• |
business combinations or significant changes in a collaborator’s business strategy may adversely affect our willingness to complete our obligations under our collaboration; or |
|
|
• |
collaborations may be terminated, which may require us to obtain additional capital to pursue further development or commercialization of the applicable product candidates. |
If our strategic collaborations do not result in the successful development and commercialization of product candidates, or if one of our collaborators terminates its agreement with us, we may not receive any future research funding or milestone or royalty payments under the collaboration or the research, development, and commercialization product that is the subject of the collaboration may be delayed. Moreover, our estimates of the potential revenue we are eligible to receive under our strategic collaborations may include potential payments related to therapeutic programs for which our collaborators have discontinued development or may discontinue development in the future. If we are unable to enter into strategic collaborations, or if any of the other events described in this paragraph occur after we enter into a collaboration, we may have to curtail the development of a particular product candidate, reduce or delay its development program or one or more of our other development programs, delay its potential commercialization or reduce the scope of our sales or marketing activities, or increase our expenditures and undertake development or commercialization activities at our own expense. If we elect to increase our expenditures to fund development or commercialization activities on our own, we may need to obtain additional capital, which may not be available to us on acceptable terms or at all. If we do not have sufficient funds, we will not be able to bring our product candidates to market and generate product revenue.
If we license products or businesses, we may not be able to realize the benefit of such transactions if we are unable to successfully integrate them with our existing operations and company culture. In addition, the success of our collaborations or other transactions may be negatively affected, including as a result of delays in timelines, if the ongoing COVID-19 pandemic materially adversely impacts our or the counterparty’s operations. We also cannot be certain that, following execution of a strategic transaction, we will achieve the revenue or specific net income that justifies such a transaction or the other anticipated benefits that led us to enter into the arrangement.
84
Our ability to develop our cell engineering platforms and product candidates and our future growth depends on retaining our key personnel and recruiting additional qualified personnel.
Our success depends upon the continued contributions of our key management, scientific, and technical personnel, many of whom have been instrumental for us and have substantial experience with our cell engineering platforms, underlying technologies, and related product candidates. Given the specialized nature of our ex vivo and in vivo cell engineering and the fact that we are operating in novel and emerging fields, there is an inherent scarcity of personnel with the requisite experience to fill the roles across our organization. As we continue developing our product candidates and building our pipeline, we will require personnel with medical, scientific, or technical qualifications specific to each program. The loss of key managers and senior scientists could delay our research and development activities. Despite our efforts to retain valuable employees, members of our management, scientific, and development teams may terminate their employment with us at any time, sometimes on short notice. Although we have employment agreements with certain of our key employees, our employment relationship with all employees provides for at-will employment, which means that any of our employees could leave our employment at any time, with or without notice. If our retention efforts are unsuccessful now or in the future, it may be difficult for us to implement our business strategy, which could have a material adverse effect on our business.
Further, certain of our key employees, including Drs. Terry Fry, Steve Goldman and Chuck Murry, retain partial employment at academic institutions. Dr. Goldman currently devotes approximately 60% of his time to the University of Rochester and the University of Copenhagen, Dr. Murry currently devotes approximately 25% to his time to the University of Washington, and Dr. Fry currently devotes approximately 25% of his time to the University of Colorado. We may in the future have other employees that have similar employment arrangements. These arrangements may expose us to increased potential for these individuals to return to their academic positions full-time or devote less of their attention to us than is optimal, and potentially expose us to claims of intellectual property ownership or co-ownership by the respective academic institutions.
The competition for qualified personnel in the biotechnology and pharmaceutical industries is intense, and our future success depends upon our ability to attract, retain, and motivate highly skilled scientific, technical, and managerial employees. Specifically, the success of our research and development programs, clinical operations, manufacturing, and future sales and marketing efforts will depend on our ability to attract and retain highly-skilled scientists, engineers, clinical operations and manufacturing personnel, and sales professionals. We face competition for personnel from other companies, universities, public and private research institutions, and other organizations. We have from time to time experienced, and we expect to continue to experience, difficulty in hiring and retaining employees with appropriate qualifications on acceptable terms, or at all. Many of the companies with which we compete for experienced personnel have greater resources than we do and may be able to provide prospective job candidates or our existing employees with more attractive roles, salaries, or benefits than we can provide. If we hire employees from competitors or other companies, their former employers may attempt to assert that these employees or we have breached legal obligations, resulting in a diversion of our time and resources and, potentially, damages. In addition, job candidates and existing employees often consider the value of the stock awards they receive in connection with their employment. If the perceived benefits of our stock awards decline or are otherwise viewed unfavorably compared to those of companies with which we compete for talent, our ability to recruit and retain highly skilled employees could be harmed. If we fail to attract new personnel or fail to retain and motivate our current personnel, our business and future growth prospects would be harmed.
Though many of our personnel have significant experience with respect to manufacturing biopharmaceutical products, we, as a company, do not have experience in developing or maintaining a manufacturing facility. We cannot guarantee that we will be able to maintain a compliant facility and manufacture our product candidates as intended, given the complexity of manufacturing novel therapeutics. If we fail to successfully operate our facility and manufacture a sufficient and compliant supply of our product candidates, our clinical trials and the commercial viability of our product candidates could be adversely affected.
The manufacture of biopharmaceutical products is complex and requires significant expertise, including the development of advanced manufacturing techniques and process controls. Manufacturers of gene and cell therapy products often encounter difficulties in production, particularly in scaling up, scaling out, validating initial production, ensuring the absence of contamination, and ensuring process robustness after initial production. These include difficulties with production costs and yields, quality control, including stability of the product, quality assurance testing, operator error, and shortages of qualified personnel, as well as compliance with strictly enforced federal, state, and foreign regulations. As a result of the complexities involved in biopharmaceutical manufacturing, the cost to manufacture biologics is generally higher than traditional small molecule chemical compounds, and the manufacturing process is less reliable and is more difficult to reproduce, and this is particularly true with respect to our product candidates. The application of new regulatory guidelines or parameters, such as those related to control strategy testing, may also adversely affect our ability to manufacture our product candidates in a compliant and cost-effective manner or at all.
We are investing early in building world class capabilities in key areas of manufacturing sciences and operations, including development of our ex vivo and in vivo cell engineering platforms, product characterization, and process analytics from the time candidates are in early research phases. Our investments also include scaled research solutions, scaled infrastructure, and novel technologies to improve efficiency, characterization, and scalability of manufacturing. However, we have limited experience in
85
managing the manufacturing processes necessary for making cell and gene therapies. We cannot be sure that the manufacturing processes that we use, or the technologies that we incorporate into these processes, will result in viable or scalable yields of ex vivo and in vivo cell engineering product candidates that will be safe and effective and meet market demand.
A key part of our strategy is operating our own manufacturing facility. Accordingly, in July 2021, we entered into a long-term lease to establish and operate our own current good manufacturing practices (cGMP) manufacturing facility to support our late-stage clinical development and early commercial product candidates across our product portfolio, including with respect to the production of allogeneic CAR T cells, viral vectors, and PSC-derived products. We expect that it will take several years before we are able to begin manufacturing our product candidates at this facility, if we are able to do so at all. Designing and building out our manufacturing facility will be time-consuming and will require significant resources, including a reallocation of certain of our existing financial, human and other resources, including the time and attention of our senior management. In addition, given the volatility in the costs of building materials, building out our manufacturing facility may be more expensive than we expect. We do not have experience as a company in developing a manufacturing facility, and we may experience unexpected costs or delays or be unsuccessful in developing our internal manufacturing capability in time to support registration-enabling clinical trials of our product candidates or at all. In order to build out the facility, we will need to engage third-party service providers and obtain equipment and third-party technology necessary to manufacture our product candidates at the facility; however, we may not be able to negotiate agreements with third parties or access necessary technologies on commercially reasonable terms or at all. Moreover, there is no guarantee that the industrial space that we are leasing to develop our manufacturing facility will not change ownership over the term of the lease or be subject to additional zoning or other restrictions, and that, in such an event, we will be able to continue to build or operate the facility without further delay or cost. In addition, operating our facility will require us to continue to hire and retain experienced scientific, quality control, quality assurance, and manufacturing personnel. As described elsewhere in these Risk Factors, competition for qualified personnel in the biotechnology and pharmaceutical industries is intense, and if we fail to attract qualified personnel or retain and motivate our current personnel, we will not be able to operate our facility, and our business and future growth prospects would be harmed.
Until we are able to begin manufacturing our product candidates at our facility, we will rely on third-party contract manufacturers to manufacture our product candidates for preclinical studies and clinical trials. Once we have completed the build-out of our manufacturing facility, we will be required to transition manufacturing processes and know-how of certain of our product candidates from our contract development and manufacturing organizations (CDMOs) to our facility. To date, we and our CDMOs have limited experience in the technology transfer of manufacturing processes from us to our CDMOs. Transferring manufacturing processes and know-how is complex and involves review and incorporation of both documented and undocumented processes that may have evolved over time. In addition, transferring production to our facility may require utilization of new or different processes to meet the requirements of our facility. Additional studies may also need to be conducted to support the transfer of certain manufacturing processes and process improvements. We will not know with certainty whether all relevant know-how and data has been adequately incorporated into the manufacturing process being conducted at our facility until the completion of studies and evaluations intended to demonstrate the comparability of material previously produced by our CDMOS with that generated by our facility.
Operating our manufacturing facility will require us to comply with complex regulations. Moreover, our manufacturing facility, and any future commercial manufacturing facilities we may operate, will require FDA or comparable foreign regulatory authority approval, which we may not obtain in time to support registration-enabling clinical trials for our product candidates, if at all. Even if approved, we would be subject to ongoing periodic unannounced inspections by the FDA, the Drug Enforcement Administration, corresponding state agencies, and comparable foreign regulatory authorities to ensure strict compliance with cGMP, current good tissue practices (cGTPs), and other government regulations. We also may make changes to our manufacturing process at various points during development, and even after commercialization, for various reasons, such as to control costs, achieve scale, decrease processing time, increase manufacturing success rate, or for other reasons. Such changes carry the risk that they will not achieve their intended objectives, and any of these changes could cause our product candidates to perform differently and affect the results of any of our then-ongoing clinical trials or future clinical trials, or the performance of the product, once commercialized. In some circumstances, changes in the manufacturing process may require us to perform comparability studies and to collect additional data from patients prior to undertaking more advanced clinical trials. For instance, changes in our process during the course of clinical development may require us to show the comparability of the product used in earlier clinical phases or at earlier portions of a trial to the product used in later clinical phases or later portions of the trial. We may also make further changes to our manufacturing process before or after commercialization, and such changes may require us to show the comparability of the resulting product to the product used in the clinical trials using earlier processes. We may be required to collect additional clinical data from any modified process prior to obtaining marketing approval for the product candidate produced with such modified process. If clinical data are not ultimately comparable to that seen in the earlier trials in terms of safety or efficacy, we may be required to make further changes to our process or undertake additional clinical testing, either of which would significantly delay the clinical development or commercialization of the relevant product candidate.
86
Furthermore, if contaminants are discovered in our supply of product candidates or in our manufacturing facility, or any future manufacturing facilities, such supply may have to be discarded and our manufacturing facilities may need to be closed for an extended period of time to investigate and remedy the contamination. We cannot guarantee that any stability or other issues relating to the manufacture of our product candidates will not occur in the future. We may not be able to manufacture our product candidates as a result of not meeting regulatory requirements and may not be able to scale up or scale out our manufacturing to meet market demand. Any failure or delay in the development of our manufacturing capabilities could adversely impact the development and potential commercialization of our product candidates.
We may encounter difficulties in managing our growth as we continue to expand our development and regulatory capabilities, which could disrupt our operations.
We have experienced rapid growth since our inception in July 2018. As of December 31, 2021, we had 383 full-time employees and three part-time employees. We expect continued growth in the number of our employees and the scope of our operations, particularly as we advance our IND-enabling studies, establish regulatory, quality, and clinical operations, and continue to establish supply chain logistics and manufacturing. To manage our anticipated future growth, we plan to continue to implement and improve our managerial, operational, and financial systems, expand our facilities, and continue to recruit and train additional qualified personnel. Due to our limited financial resources and the complexity involved in managing a company with such anticipated growth, we may not be able to effectively manage the expansion of our operations or recruit and train additional qualified personnel. In addition, we have limited experience in managing the manufacturing processes necessary for making cell and gene therapies. The expansion of our operations will be costly and may divert our management and business development resources. For example, members of management will have significant added responsibilities in connection with effecting and managing our growth, including identifying, recruiting, integrating, maintaining, and motivating current and future employees, effectively managing our internal development efforts, including the clinical and regulatory (e.g., FDA) review process, while complying with our contractual obligations to third parties, and maintaining and improving our operational, financial, and management controls, reporting systems, and procedures. In addition, as we grow, we may be required to rely more heavily on third-party service providers, advisors, and consultants to provide certain services, including strategic, financial, business development, and research and development services, as well as with respect to certain aspects of our regulatory approval affairs and manufacturing activities. We cannot guarantee that such third parties will be available to us on a timely basis when needed, or that we will be able to find and engage qualified replacements if required. Our inability to successfully manage our growth could delay the execution of our business plans or disrupt our operations.
We may expend our limited resources to pursue a particular product candidate or indication and fail to capitalize on product candidates or indications that may be more profitable or for which there is a greater likelihood of success.
Because we have limited financial and managerial resources, we focus on research programs, therapeutic platforms, and product candidates that we identify for specific indications. Additionally, we have contractual commitments under our collaboration agreements to use commercially reasonable efforts to develop certain programs and, thus, do not have unilateral discretion to vary from such agreed upon efforts. In addition, we have contractual commitments to conduct certain development plans, and thus may not have discretion to modify such development plans, including clinical trial designs, without agreement from our collaboration partners. As a result, we may forego or delay pursuit of opportunities with other therapeutic platforms or product candidates or for other indications that later prove to have greater commercial potential. Our resource allocation decisions may cause us to fail to capitalize on viable commercial products or profitable market opportunities. Our spending on current and future research and development programs, therapeutic platforms, and product candidates for specific indications may not yield any commercially viable products. If we do not accurately evaluate the commercial potential or target market for a particular product candidate, we may relinquish valuable rights to that product candidate through collaboration, licensing, or other royalty arrangements in cases in which it would have been more advantageous for us to retain sole development and commercialization rights.
The use of human stem cells exposes us to a number of risks in the development of our human stem cell-derived products, including an inability to obtain suitable donor material from eligible and qualified human donors, restrictions on the use of human stem cells, as well as ethical, legal, and social implications of research on the use of stem cells, any of which could prevent us from completing the development of or commercializing and gaining acceptance for our products derived from human stem cells.
We use human stem cells in our research and development, including induced PSCs (iPSCs) and embryonic stem cells (ESCs), and one or more of our ex vivo cell engineering product candidates may be derived from human stem cells. The use of such cells in our research, or as starting cell lines in the manufacture of one or more of our product candidates, exposes us to a number of risks. These risks include difficulties in securing sufficient and viable stem cells as starting material, recruiting patients for our future clinical trials, as well as managing a multitude of legal and regulatory restrictions on the sourcing and use of these cells. In particular, in some states, use of embryonic tissue as a source of stem cells is prohibited and many research institutions have adopted policies regarding the ethical use of human embryonic tissue. If these policies or restrictions have the effect of limiting the scope of research we can conduct using stem cells, our ability to develop our ex vivo cell engineering product candidates may be significantly impaired, which could
87
have an adverse material effect on our business. Further, the use of stem cells generally, and embryonic stem cells in particular, has social, legal, and ethical implications. Certain political and religious groups continue to voice opposition to the use of human stem cells in drug research, development, and manufacture. Adverse publicity due to ethical and social controversies surrounding the use of stem cells could lead to negative public opinion, difficulties enrolling patients in our clinical trials, increased regulation, and stricter policies regarding the use of such cells, which could harm our business and may limit market acceptance of any of our product candidates that may receive regulatory approval. In addition, clinical experience with stem cells, including iPSCs and ESCs, is limited. We are not aware of any products that utilize iPSCs or ESCs as a starting material that have received marketing approval from the FDA or a comparable foreign regulatory authority. Therefore, patients in our clinical trials may experience unexpected side effects, and we may experience unexpected regulatory delays prior to or after regulatory approval, if approval were to be granted.
Furthermore, manufacturing and development of our ex vivo stem cell-derived and allogeneic T cell-derived product candidates will rely on our ability to obtain suitable donor material from eligible and qualified human donors. If we are unable to obtain sufficient quantities of suitable donor material, or if we are unable to obtain such material in a timely manner, we may experience delays in manufacturing our ex vivo product candidates, which would harm our ability to conduct future clinical trials for or to commercialize these product candidates. Moreover, if the consent, authorization, or process for the donation of those materials is not obtained or conducted in accordance with applicable legal, ethical, and regulatory requirements, we could face delays in the clinical testing and approval of these product candidates, or, potentially, we could face claims by such human donors, which could expose us to damages and reputational harm.
The ongoing COVID-19 pandemic could materially and adversely affect our preclinical studies and development, our manufacturing capabilities, any clinical trials we may commence, and our business, financial condition, and results of operations.
As a result of the COVID-19 pandemic, or similar pandemics, and related “shelter in place” orders and other public health guidance measures, we have experienced and may in the future experience disruptions that could materially and adversely impact our preclinical studies and development, any clinical trials we may commence, and our business, financial condition, and results of operations. In response to the spread of COVID-19, we have limited operations in our executive offices, with our administrative employees primarily continuing their work outside of our offices, and have taken other precautionary measures, including the periodic testing of our on-site employees. We also established a cross-functional task force and implemented business continuity plans designed to address and mitigate the impact of the ongoing COVID-19 pandemic on our business.
Potential disruptions to our preclinical development efforts resulting from the ongoing COVID-19 pandemic may include the following:
|
|
• |
delays or disruptions in preclinical experiments and IND-enabling studies due to restrictions of on-site staff, limited or no access to animal facilities, and unforeseen circumstances at contract research organizations (CROs) and vendors; |
|
|
• |
limitations on employee or other resources that would otherwise be focused on the conduct of our preclinical activities, including because of illness of employees or their families, the desire of employees to avoid travel or contact with large groups of people, an increased reliance on working from home, school closures, or mass transit disruptions; |
|
|
• |
delays in necessary interactions with regulatory authorities, ethics committees, and other important agencies and contractors due to limitations in employee resources or forced furlough of government or contractor personnel; and |
|
|
• |
limitations in maintaining our corporate culture that facilitates the transfer of institutional knowledge within our organization and fosters innovation, teamwork, and a focus on execution. |
In addition, we have experienced, and we and our service providers or vendors may continue to experience, delays in the procurement of, or an inability to procure, certain laboratory supplies required for the conduct of our research and preclinical activities, such as cell culture plasticware and single use containers, as a result of factors related to the ongoing COVID-19 pandemic, including increased demand due to ramp up of COVID-19 research and manufacturing, government-mandated allocation of materials for such research and manufacturing, insufficient manufacturing capacity, and delays by CDMOs in increasing manufacturing capacity to address increased demand. The ongoing COVID-19 pandemic may also adversely affect our manufacturing capabilities. For example, we may experience delays or otherwise experience difficulties in building out and operationalizing our internal manufacturing facility and obtaining key materials, consumables, and equipment necessary to manufacture our product candidates.
In addition, if and when we commence clinical trials for any of our product candidates, we may experience potential delays or disruptions of clinical trial-related activities as a result of the ongoing COVID-19 pandemic, including as a result of the following:
|
|
• |
interruption of key clinical trial activities, such as clinical trial site data monitoring and efficacy, safety, and translational data collection, processing, and analyses, due to limitations on travel imposed or recommended by federal, state, or local governments, employers, and others, or interruption of clinical trial subject visits, which may impact the collection and integrity of subject data and clinical study endpoints; |
88
|
|
• |
delays or difficulties in initiating or expanding clinical trials, including delays or difficulties with clinical site initiation and recruiting clinical site investigators and staff; |
|
|
• |
delays or difficulties in enrolling and retaining patients in our clinical trials; |
|
|
• |
increased rates of patient withdrawal from our clinical trials following enrollment as a result of contracting COVID-19, developing other health conditions, or being forced to quarantine; |
|
|
• |
interruption of, or delays in receiving, supplies of our product candidates from our contract manufacturing organizations due to staffing shortages, production slowdowns, or stoppages and disruptions in delivery systems with respect to materials and reagents; |
|
|
• |
diversion of healthcare resources away from the conduct of our clinical trials toward efforts to support the COVID-19 pandemic response, including the diversion of resources, including staff, at hospitals serving as our clinical trial sites and supporting our clinical trials; |
|
|
• |
interruption or delays in the operations of the FDA and comparable foreign regulatory agencies; |
|
|
• |
changes in regulations implemented in response to the COVID-19 pandemic that may require us to change the ways in which our clinical trials are conducted, which may result in unexpected costs, or to discontinue the clinical trials altogether; |
|
|
• |
delays in receiving approval from local regulatory authorities to initiate our planned clinical trials; |
|
|
• |
limitations on employee resources that would otherwise be focused on the conduct of our clinical trial-related activities, including because of illness of employees or their families or the desire of employees to avoid contact with large groups of people; and |
|
|
• |
additional delays, difficulties, or interruptions as a result of current or future shutdowns or other restrictions imposed in response to the COVID-19 pandemic in countries where we or our third-party service providers operate. |
The COVID-19 global pandemic continues to rapidly evolve. Although many countries, including certain countries in Europe and the United States, have re-opened, rises in new cases, including as the result of newly identified COVID-19 variants, have caused certain countries, states, and localities to re-initiate restrictions. The extent to which the COVID-19 pandemic may affect our preclinical studies, future clinical trials, business, financial condition, and results of operations will depend on future developments, which are highly uncertain and cannot be predicted at this time, such as the geographic spread of the disease, the duration of the pandemic, travel restrictions, actions to contain the pandemic or reduce its impact in the United States and other countries, such as required social distancing, quarantines, lock-downs, business closures, or business disruptions, and the effectiveness of actions taken in the United States and other countries to contain and treat the disease. Additionally, we are unable to predict if and when a different pandemic may occur, and if so, whether it would have similar or different impacts on our business, financial condition, or share price. Future developments in these and other areas present material uncertainty and risk with respect to our preclinical activities, clinical trials, business, financial condition, and results of operations.
Negative public opinion and increased regulatory scrutiny of research and therapies involving gene editing or other ex vivo or in vivo cell engineering technologies may damage public perception of our product candidates or adversely affect our ability to conduct our business or obtain regulatory approvals for our product candidates.
Certain aspects of our cell engineering platforms rely on the ability to edit genes. Public perception may be influenced by claims that gene editing is unsafe, and products incorporating gene editing may not gain the acceptance of the public or the medical community. In particular, our success will depend upon physicians specializing in diseases that our product candidates are designed to target prescribing our product candidates as treatments in lieu of, or in addition to, existing, more familiar treatments for which greater clinical data may be available. Any increase in negative perceptions of gene editing may result in fewer physicians prescribing our treatments or may reduce the willingness of patients to utilize our treatments or participate in clinical trials of our product candidates. In addition, given the novel nature of ex vivo and in vivo cell engineering technologies, governments may impose import, export, or other restrictions in order to retain control or limit the use of such technologies. Increased negative public opinion or more restrictive government regulations, either in the United States or internationally, would have a negative effect on our business or financial condition and may delay or impair the development and commercialization of our product candidates or demand for such product candidates.
89
Risks Related to the Development and Clinical Testing of Our Product Candidates
We must successfully progress our product candidates through extensive preclinical studies and clinical trials in order to obtain regulatory approval to market and sell such product candidates. Even if we obtain positive results in preclinical studies of a product candidate, these results may not be predictive of the results of future preclinical studies or clinical trials.
To obtain the requisite regulatory approvals to market and sell any of our product candidates, we or any future collaborator for such product candidate must demonstrate through extensive preclinical studies and clinical trials that the product candidate is safe, pure, and potent in humans. Before an IND can be submitted to the FDA and become effective, which is a prerequisite for conducting clinical trials on human subjects, a product candidate must successfully progress through extensive preclinical studies, which include preclinical laboratory testing, animal studies, and formulation studies conducted in accordance with good laboratory practices (GLP).
Success in preclinical studies does not ensure that later preclinical studies or clinical trials will be successful. A number of companies in the biotechnology and pharmaceutical industries have suffered significant setbacks in clinical trials, even after positive results in preclinical studies. These setbacks have been caused by, among other things, preclinical findings made while clinical trials were underway and safety or efficacy observations made during the course of clinical trials, including previously unreported adverse events. The design of a clinical trial can determine whether its results will support approval of a product, and flaws in the design of a clinical trial may not become apparent until the clinical trial is well advanced. In addition, preclinical and clinical data are often susceptible to varying interpretations and analyses. Notwithstanding any potential promising results in earlier studies, we cannot be certain that we will not face similar setbacks. In addition, the results of our preclinical animal studies, including our non-human primate studies, may not be predictive of the results of subsequent clinical trials on human subjects. Product candidates may fail to show the desired pharmacological properties or safety and efficacy traits in clinical trials despite having successfully progressed through preclinical studies.
If we fail to obtain positive results in preclinical studies or clinical trials of any product candidate, the development timeline and regulatory approval and commercialization prospects for that product candidate, and, correspondingly, our business and financial prospects, would be negatively impacted.
All of our product candidates are in preclinical development, and none have commenced clinical development. Preclinical and clinical drug development is a lengthy and expensive process with uncertain timelines and uncertain outcomes. If preclinical studies or clinical trials of any of our product candidates are prolonged or delayed, we may be unable to obtain required regulatory approvals and commercialize such product candidates on a timely basis or at all.
Preclinical studies and clinical trials are expensive, can take many years to complete, and their outcomes are inherently uncertain. Failure can occur at any time during preclinical or clinical development. Product candidates in later-stage clinical trials may fail to produce the same results as observed in earlier trials or fail to show the desired safety and efficacy traits despite having progressed through preclinical studies and earlier clinical trials. Our future clinical trials may not be successful.
Applicable laws and regulations require us to test our product candidates in animals before initiating clinical trials involving humans. We may experience delays or experience difficulty completing studies of our product candidates in animals for various reasons. For example, due to global supply chain issues caused by global geo-political, economic, and other factors beyond our control, including the ongoing COVID-19 pandemic, as described elsewhere in these Risk Factors, we have experienced and may continue to experience difficulty in accessing animal models, specifically non-human primate models, for the preclinical evaluation of our product candidates. In addition, animal testing activities have been the subject of controversy and adverse publicity. Animal rights groups and other organizations and individuals have attempted to stop animal testing activities by pressing for legislation and regulation in these areas and by disrupting these activities through protests and other means. To the extent the activities of these groups are successful, our research and development activities may be interrupted, delayed, or become more expensive.
We are required to submit an IND to the FDA with respect to each product candidate prior to commencing a clinical trial for such product candidate. While we plan to submit INDs for each of our product candidates, we may not be able to submit such INDs in accordance with our expected timelines for various reasons, including due to:
|
|
• |
manufacturing delays, including due to challenges associated with scaling up our manufacturing processes and developing and validating assays; |
|
|
• |
delays with IND-enabling preclinical studies; or |
|
|
• |
feedback from the FDA that requires us to conduct additional testing or change the design of a planned clinical trial prior to submitting such IND. |
Moreover, we cannot guarantee that submission of an IND for a product candidate will result in the FDA or comparable foreign regulatory authorities allowing clinical trials of that product candidate to commence in accordance with our timelines or expectations
90
or at all. For example, the FDA may accept an IND submission for a product candidate but place clinical trials of such product candidate on hold pending the results of additional testing or the development of additional assays. Additionally, even if regulatory authorities agree with the design and implementation of the clinical trials set forth in an IND, we cannot guarantee that such regulatory authorities will not change their requirements in the future. These considerations also apply to new clinical trials we may submit as amendments to existing INDs.
To date, we have not commenced any clinical trials. We do not know whether planned clinical trials will begin on time, need to be redesigned, enroll patients on time, or be completed on schedule, if at all. Clinical trials may be delayed, suspended, or terminated for a variety of reasons, including the following:
|
|
• |
delays in or failure to obtain regulatory authorization to commence a trial; |
|
|
• |
delays in or failure to obtain institutional review board (IRB) approval at each clinical trial site; |
|
|
• |
delays in or failure to reach agreement with prospective CROs and clinical trial sites on acceptable terms, or at all, which agreements can be subject to extensive negotiation and may vary significantly among different CROs and trial sites; |
|
|
• |
difficulty in recruiting clinical trial investigators of appropriate competencies and experience; |
|
|
• |
lack of sufficient availability of suitable donor material from eligible and qualified donors for the manufacture of product candidates from our ex vivo cell engineering platform; |
|
|
• |
delays in establishing the appropriate dosage levels in clinical trials; |
|
|
• |
delays in or inability to recruit and enroll suitable patients to participate in a trial, including as a result of study inclusion and exclusion criteria and patients’ prior lines of therapy and treatment; |
|
|
• |
the difficulty in certain countries in identifying the sub-populations that are the target group for a particular trial, which may delay enrollment and reduce the power of a clinical trial to detect statistically significant results; |
|
|
• |
lower than anticipated retention rates of patients in clinical trials; |
|
|
• |
failure of patients to complete a trial or return for post-treatment follow-up; |
|
|
• |
clinical sites deviating from trial protocol or dropping out of a trial; |
|
|
• |
delays caused by the addition of new investigators or clinical trial sites; |
|
|
• |
safety or tolerability concerns relating to the product candidate being tested that could cause us or governmental authorities, as applicable, to suspend or terminate a clinical trial, including if participants are being exposed to unacceptable health risks or experiencing undesirable side effects or there are other unfavorable characteristics of the product candidate, or if there is evidence that potential undesirable side effects or risks may be associated with a therapeutic or therapeutic candidate being developed by us or a third party and regulators deem our product candidate to have the potential for comparable side effects or risks as such therapeutic or therapeutic candidate because of biologic, mechanistic, sourcing, or other similarities; |
|
|
• |
the failure of third-party research contractors to comply with regulatory requirements or meet their contractual obligations in a timely manner or at all; |
|
|
• |
changes in regulatory requirements, policies, and guidelines; |
|
|
• |
inability to manufacture sufficient quantities of a product candidate for use in clinical trials; |
|
|
• |
the quality or stability of a product candidate falling below acceptable standards, or failure to manufacture product candidates in accordance with cGMP and other applicable laws, regulations and guidelines; |
|
|
• |
changes in the treatment landscape for our target indications that may make our product candidates no longer relevant; |
|
|
• |
claims that the product candidate being tested infringes third-party intellectual property rights, including any resulting injunctions that may prevent further use of such product candidates and interfere with the progress of the trial; and |
|
|
• |
business interruptions resulting from geo-political actions, including war and terrorism, natural disasters including earthquakes, typhoons, floods, and fires, or disease, including the ongoing COVID-19 pandemic. |
In addition, disruptions caused by the COVID-19 pandemic, to the extent it is still ongoing when we initiate our planned clinical trials, may increase the likelihood that we encounter difficulties or delays in initiating, enrolling, conducting, or completing such clinical trials, as described elsewhere in these Risk Factors.
91
Additionally, some of our trials may be open-label trials in which both the patient and investigator know whether the patient is receiving the investigational product candidate or an existing approved therapy. Open-label clinical trials are subject to various limitations that may exaggerate any therapeutic effect, as patients in open-label clinical trials are aware when they are receiving treatment. In addition, open-label clinical trials may be subject to an “investigator bias,” where those assessing and reviewing the physiological outcomes of patients in the clinical trials are aware of which patients have received the experimental treatment and may interpret the information of this group more favorably given this knowledge. Therefore, it is possible that positive results observed in open-label trials will not be replicated in later placebo-controlled trials.
Clinical trials must be conducted in accordance with the FDA and comparable foreign regulatory authorities’ legal requirements, regulations and guidelines and are subject to oversight by these governmental authorities and IRBs or Ethics Committees at the medical institutions where the clinical trials are conducted. We could encounter delays if a clinical trial is suspended or terminated by us, by the IRBs or Ethics Committees of the institutions at which such trial is being conducted, by the Data Review Committee or Data Safety Monitoring Board for such trial, or by the FDA or comparable foreign regulatory authorities. Such authorities may impose such a suspension or termination, including following an inspection of clinical trial operations or a clinical trial site, due to a number of factors, including failure to conduct the clinical trial in accordance with regulatory requirements or our clinical protocols, unforeseen safety issues or adverse side effects, failure to demonstrate a benefit from use of the product candidate being tested, changes in governmental regulations or administrative actions, or lack of adequate funding to continue the clinical trial. In addition, such authorities may impose a clinical hold on a product candidate due to unforeseen safety issues or adverse side effects that may be associated with a therapeutic or therapeutic candidate being developed by us or a third party if regulators deem our product candidate to have the potential for comparable side effects or risks as such therapeutic or therapeutic candidate because of biologic, mechanistic, sourcing, or other similarities. If we experience delays in completing, or are required to terminate, any clinical trial of our product candidates, the commercial prospects of the relevant product candidates will be harmed, and our ability to generate product revenues from these product candidates will be delayed. In addition, any delays in completing our clinical trials will increase our costs, delay our ability to obtain regulatory approval for the relevant product candidate, and jeopardize our ability to commence product sales and generate revenues. Significant clinical trial delays could also allow our competitors to bring products to market before we do or shorten any periods during which we have the exclusive right to commercialize our product candidates, which may impair our ability to commercialize our product candidates and harm our business and results of operations.
Furthermore, as described elsewhere in these Risk Factors, we will rely on CROs and clinical trial sites to ensure the proper and timely conduct of our clinical trials in compliance with good clinical practices (GCP) requirements. While we will enter into agreements governing their conduct, we will have limited influence over their actual performance. To the extent the CROs and clinical trial sites fail to timely and successfully enroll patients in our clinical trials, fail to conduct such clinical trials in accordance with GCP, or experience significant delays in the execution of trials, including delays in achieving full enrollment or clinical trial data collection and analysis, we may experience program delays, incur additional costs, or both, which may harm our business. In addition, we may experience delays and incur additional costs with respect to clinical trials that we conduct in countries outside the United States, including as a result of increased shipment and distribution costs, compliance with additional regulatory requirements, and the engagement of non-United States. CROs, and may also be exposed to risks associated with clinical investigators who are unknown to the FDA, and different standards of diagnosis, screening, and medical care.
Our future clinical trials may fail to demonstrate substantial evidence of the safety and efficacy of our product candidates, including any future product candidates, which would prevent, delay or limit the scope of regulatory approval and commercialization of such product candidates.
To obtain the requisite regulatory approvals to market and sell any of our product candidates and any other future product candidates, we must demonstrate through clinical trials that our product candidates are safe and effective for use in each targeted indication. Clinical testing is expensive and can take many years to complete, and its outcome is inherently uncertain. Failure can occur at any time during the clinical development process. Most product candidates that begin clinical trials are never approved by regulatory authorities for commercialization. We may be unable to establish clinical endpoints that applicable regulatory authorities would consider clinically meaningful.
Further, the process of obtaining regulatory approval is expensive, often takes many years following the commencement of clinical trials, and can vary substantially based upon the type, complexity, and novelty of the product candidates involved, as well as the target indications, patient population, and regulatory authority involved. Prior to obtaining approval to commercialize our current or future product candidates in the United States or abroad, we or our potential future collaborators must demonstrate with substantial evidence from adequate and well-controlled clinical trials, and to the satisfaction of the FDA or comparable foreign regulatory authorities, that such product candidates are safe and effective for their intended uses.
Clinical trials that we conduct may not demonstrate the efficacy and safety necessary to obtain regulatory approval to market our product candidates. In some instances, there can be significant variability in safety or efficacy results between different clinical trials of the same product candidate due to numerous factors, including changes in trial procedures set forth in protocols, differences in
92
the size and type of the patient populations, changes in and adherence to the clinical trial protocols, and the rate of dropout among clinical trial participants. If the results of our clinical trials are inconclusive with respect to the efficacy of our product candidates, if we do not meet the clinical endpoints with statistical and clinically meaningful significance, or if there are safety concerns associated with our product candidates, we may experience delays in obtaining marketing approval, or we may not obtain approval at all. Additionally, any safety concerns observed in any one of our clinical trials for a product candidate in our targeted indications could limit the prospects for regulatory approval of such product candidate in those and other indications.
Even if we successfully complete any future clinical trials, clinical data are often susceptible to varying interpretations and analyses. We cannot guarantee that the FDA or comparable foreign regulatory authorities will interpret the results as we do, and more trials could be required before we submit our product candidates for approval. Even if positive results are observed in clinical trials, we cannot guarantee that the FDA or comparable foreign regulatory authorities will view our product candidates as having efficacy. Further, the FDA or comparable foreign regulatory authorities may not agree with our manufacturing strategy or may not find comparability between our clinical trial product candidates and proposed commercial product candidates, which may result in regulatory delays or a need to perform additional clinical studies. Moreover, clinical trial results that may be acceptable to support approval of a certain scope in one jurisdiction may be deemed inadequate to support regulatory approval, or may only be deemed sufficient to support a narrower scope of approval, in other jurisdictions. If the FDA or comparable foreign regulatory authorities determine that our clinical trial results are not adequate to support approval of a marketing application, we may experience delays in obtaining, or fail to obtain, approval of our product candidates, or we may be required to expend significant additional resources, which may not be available to us, to conduct additional trials in support of potential approval of our product candidates. Even if regulatory approval is obtained for a product candidate, the terms of such approval may limit the scope and use of the specific product candidate, which may also limit its commercial potential.
Our product candidates may have serious adverse, undesirable, or unacceptable side effects or other properties that may delay or prevent marketing approval. If a product candidate receives regulatory approval, and such side effects are identified following such approval, the commercial profile of any approved label may be limited, or we may be subject to other significant negative consequences following such approval.
Our product candidates may cause undesirable side effects, which could cause us or regulatory authorities to interrupt, delay, or halt our future clinical trials and could result in a more restrictive label or the delay or denial of regulatory approval by the FDA or comparable foreign authorities. We have not commenced clinical trials for any of our product candidates, and we do not have any clinical data or other information that would enable us to fully anticipate their side effects. Accordingly, we may observe unexpected side effects or higher levels of known side effects in clinical trials of our product candidates, including adverse events known to occur in the same classes of therapeutics. These may include, among others, infusion reaction, cytokine release syndrome (CRS), graft-versus-host disease (GvHD), neurotoxicities, and certain cancers.
Results of our clinical trials could reveal a high and unacceptable severity and prevalence of these or other side effects associated with our product candidates. In such an event, clinical trials of such product candidates could be suspended or terminated, and the FDA or comparable foreign regulatory authorities could order us to cease further development of or deny approval of such product candidates for any or all targeted indications. The occurrence of such side effects could negatively affect our ability to recruit and enroll patients in our clinical trials, or the ability of enrolled patients to complete the clinical trials or result in product liability claims. Any of these occurrences may harm our business, financial condition, and prospects significantly.
Further, clinical trials by their nature utilize only a sample of the potential patient population. With a limited number of patients and limited duration of exposure to our product candidates, rare and severe side effects of our product candidates may not be apparent during early clinical trials and may only be uncovered once a significantly larger number of patients have been exposed to the product candidate, including during later-stage clinical trials or following commercialization.
In the event that any of our product candidates receives marketing approval and we or others later identify undesirable or unacceptable side effects caused by such products, a number of potentially significant negative consequences could result, including:
|
|
• |
regulatory authorities may withdraw or limit approvals of such products and require us to take such products off the market; |
|
|
• |
regulatory authorities may require the addition of labeling statements, specific warnings, or a contraindication or field alerts to physicians and pharmacies, or issue other communications containing warnings or other safety information about the product; |
93
|
|
• |
regulatory authorities may require a medication guide outlining the risks of such side effects for distribution to patients or that we implement a risk evaluation and mitigation strategy (REMS) plan to ensure that the benefits of the product outweigh its risks; |
|
|
• |
we may be required to change the therapeutic dose or the way the product is administered, conduct additional clinical trials, or change the labeling of the product; |
|
|
• |
we may be subject to limitations on how we may promote or manufacture the product; |
|
|
• |
sales of the product may decrease significantly; |
|
|
• |
we may be subject to litigation or product liability claims; and |
|
|
• |
our reputation may suffer. |
Any of these events could prevent us or our potential future partners from achieving or maintaining market acceptance of the affected product or could substantially increase commercialization costs and expenses, which in turn could delay or prevent us from generating significant revenue from the sale of any products.
Interim, topline, or preliminary data from our preclinical studies or future clinical trials that we may announce or publish from time to time may change as more patient data become available or as we make changes to our manufacturing processes. These data are subject to audit and verification procedures that could result in material changes in the final data.
From time to time, we may publicly disclose interim, topline, or preliminary data from our preclinical studies or future clinical trials, which are based on a preliminary analysis of then-available data, and the results and related findings and conclusions are subject to change following a more comprehensive review of the data related to the particular study or trial. We also make assumptions, estimations, calculations, and conclusions as part of our analyses of data, and we may not have received or had the opportunity to fully and carefully evaluate all data at the time of our initial disclosure of data. Further, modifications or improvements to our manufacturing processes for a product candidate may result in changes to its characteristics or behavior that could cause the product candidate to perform differently and affect the results of our ongoing clinical trials of such product candidate. As a result, the topline results that we report may differ from future results of the same studies, or different conclusions or considerations may qualify such results once additional data have been received and fully evaluated. Topline data also remain subject to audit and verification procedures that may result in the final data being materially different from the preliminary data we previously disclosed. As a result, topline data should be viewed with caution until the final data are available. Similarly, preliminary or interim data from clinical trials are subject to the risk that one or more of the clinical outcomes may materially change as patient enrollment continues and more patient data become available. Adverse differences between preliminary or interim data and final data could significantly harm our business prospects. Additionally, disclosure of preliminary or interim data by us or our competitors, with respect to clinical trials of their product candidates, could result in volatility in the price of our common stock.
Further, others, including regulatory authorities, may not accept or agree with our assumptions, estimates, calculations, conclusions, or analyses or may interpret or weigh the importance of data differently, which could impact the value of the particular program, the approvability or commercialization of the particular product candidate, and our company in general. If the interim, topline, or preliminary data that we report differ from actual results, or if others, including regulatory authorities, disagree with the conclusions reached, our ability to obtain approval for and commercialize our potential product candidates may be harmed, which could harm our business, operating results, prospects, or financial condition.
The manufacture of our product candidates is complex. We or our third-party CDMOs may encounter difficulties in production, which could delay or entirely halt our or their ability to supply our product candidates for clinical trials or, if approved, for commercial sale.
Our product candidates are considered to be biologics, and the process of manufacturing biologics is complex and requires significant expertise and capital investment, including the development of advanced manufacturing techniques and process controls. In July 2021, we entered into a long-term lease to establish and operate our own cGMP manufacturing facility to support our late-stage clinical development and early commercial product candidates across our product portfolio. However, we expect that it will take several years before we are able to begin manufacturing our product candidates at this facility, if at all. We currently rely, and expect for some period of time to continue to rely, on third-party CDMOs for the manufacture of our product candidates for preclinical and clinical studies. To date, we and our CDMOs have limited experience in manufacturing of cGMP batches of our product candidates. Our CDMOs and, once we begin to operate our manufacturing facility, we, must comply with cGMPs and other regulations and guidelines applicable to the manufacturing of biologics for use in clinical trials and, if approved, commercial sale. To date, we have not scaled the manufacturing processes with respect to our product candidates for later-stage clinical trials and commercialization. Larger-scale manufacturing will require the development of new processes, including for the removal of impurities that are a normal byproduct of the manufacturing process. The nature of our product candidates requires the development of novel manufacturing
94
processes and analytical technologies, which could cause delays in the scaling of manufacturing, as well as greater costs that could negatively impact the financial viability of our product candidates. We cannot be sure that the manufacturing processes employed by our CDMOs or the technologies that our CDMOs incorporate into our manufacturing processes will result in viable or scalable yields of ex vivo and in vivo cell engineering product candidates that will be safe and effective and, if approved, meet market demand.
Once we have completed the build-out of our manufacturing facility, we will be required to transition manufacturing processes and know-how of certain of our product candidates from our CDMOs to our facility. To date, we and our CDMOs have limited experience in the technology transfer of manufacturing processes from us to our CDMOs. Transferring manufacturing processes and know-how is complex and involves review and incorporation of both documented and undocumented processes that may have evolved over time. In addition, transferring production to our facility may require utilization of new or different processes to meet the requirements of our facility. Additional studies may also need to be conducted to support the transfer of certain manufacturing processes and process improvements. We will not know with certainty whether all relevant know-how and data has been adequately incorporated into the manufacturing process being conducted at our facility until the completion of studies and evaluations intended to demonstrate the comparability of material previously produced by our CDMOs with that generated by our facility.
The process of manufacturing our biologic product candidates is extremely susceptible to product loss due to contamination, equipment failure, or improper installation or operation of equipment, vendor or operator error, inconsistency in yields, variability in product characteristics, and difficulties in scaling the production process. Even minor deviations from normal manufacturing processes could result in reduced production yields, product defects, and other supply disruptions. If microbial, viral, or other contaminations are discovered in our product candidates or in the manufacturing facilities in which our product candidates are made, this could lead to withdrawal of our products from clinical trials and, if approved, the market, and such manufacturing facilities may need to be closed for an extended period of time to investigate and remedy the contamination. Moreover, if the FDA or comparable foreign regulatory authorities determine that we or our CDMOs are not in compliance with applicable laws and regulations, including those governing cGMPs, the FDA or comparable foreign regulatory authority may not approve a BLA or comparable foreign marketing authorization until the deficiencies are corrected or we replace the manufacturer in our applications with a manufacturer that is in compliance. If we or our CDMOs fail to comply with applicable regulatory requirements, we may ultimately be unable to manufacture our product candidates.
Any adverse developments affecting manufacturing operations for any of our product candidates for which we may obtain approval, may result in shipment delays, inventory shortages, lot failures, product withdrawals or recalls, or other supply interruptions. We may also have to take inventory write-offs and incur other charges and expenses for products that fail to meet specifications as a result of defects or storage over an extended period of time, undertake costly remediation efforts, or seek more costly manufacturing alternatives. As part of our process development efforts, we also may make changes to our manufacturing processes at various points during development for various reasons, such as to control costs, achieve scale, decrease processing time, increase manufacturing success rate, or other reasons. Such changes may not achieve their intended objectives, and any of these changes could cause our product candidates to perform differently and affect the results of our future clinical trials. In some circumstances, changes in the manufacturing process for a product candidate may require us to perform comparability studies and collect additional data from patients prior to undertaking more advanced clinical trials.
Given the complexities associated with manufacturing our product candidates, our ability to successfully conduct clinical trials and ultimately commercialize our product candidates will depend in part on our ability to attract, motivate, and retain highly-skilled personnel with significant expertise in manufacturing biologics that can effectively and timely manage and conduct our manufacturing operations. We and our CDMOs face intense competition to attract, motivate, and retain qualified personnel. If we or our CDMOs are unable to attract, motivate, and retain qualified personnel to conduct and manage our manufacturing operations, we may experience delays in manufacturing our product candidates, which could materially harm our ability to conduct our clinical trials or commercialize our product candidates in a timely manner or at all and could harm our business.
We are exposed to a number of risks related to the supply chain for the materials required to manufacture our product candidates.
Manufacturing our product candidates is highly complex and requires sourcing of specialty materials. Many of the risks associated with the complexity of manufacturing our final product candidates are applicable to the manufacture and supply of the raw materials required to make such product candidates. In particular, these raw materials are subject to inconsistency in yields, variability in characteristics, contamination, difficulties in scaling the production process, and defects. Similar minor deviations in the manufacturing process for these raw materials could result in supply disruption and reduced production yields for our final product candidates. In addition, we rely on third parties for the supply of these materials, which, exposes us to risks associated with dependence on third parties, as described elsewhere in these Risk Factors.
We rely on obtaining suitable donor material from eligible and qualified donors for the manufacture of product candidates from our ex vivo cell engineering platform. We may not be able to obtain sufficient quantities donor material in a timely manner or at all, including due to an inability to find donors who meet the eligibility criteria or geo-political, economic, and other factors beyond our control, including the ongoing COVID-19 pandemic, that may prevent individuals from donating blood. If we are unable to obtain
95
sufficient quantities of suitable donor material, or if we are unable to obtain such material in a timely manner, we may experience delays in manufacturing our ex vivo product candidates, which would harm our ability to conduct future clinical trials for or to commercialize these product candidates.
In addition, our manufacturing processes require many reagents, which are drug substance intermediates used in our manufacturing processes to bring about chemical or biological reactions, and other specialty materials, consumables, and equipment, some of which are manufactured or supplied by small companies with limited resources and experience with respect to supporting clinical or commercial biologics production. We currently depend on a limited number of vendors for certain materials and equipment used in the manufacture of our product candidates. Some of these suppliers may not have the capacity to support manufacturing of products under cGMP or may otherwise be ill-equipped to support our needs. Reagents and other key materials from these suppliers may have inconsistent attributes and introduce variability into our manufactured product candidates, which may contribute to variable patient outcomes and possible adverse events. We also do not have supply contracts with many of these suppliers and may not be able to enter into supply contracts with them on acceptable terms or at all. Accordingly, we may experience delays in receiving key materials and equipment to support clinical or commercial manufacturing.
For some of these reagents, materials, and equipment, we rely and may in the future rely on sole source vendors or a limited number of vendors. We may be unable to continue to source reagents, materials, or equipment from any of these suppliers for various reasons, including due to regulatory actions or requirements affecting a supplier, adverse financial or other strategic developments experienced by a supplier, labor disputes or shortages, unexpected demands, or quality issues. Additionally, due to global geo-political, economic, and other factors beyond our control, including the ongoing COVID-19 pandemic, there has been, and there may continue to be, a shortage of key materials and equipment that are necessary to manufacture our product candidates, including certain consumables such as bags, flasks, and pipette tips, which could affect our or our contract manufacturers’ ability to obtain the materials and equipment necessary to manufacture our product candidates. If any of the foregoing events were to occur, we may experience delays in manufacturing our product candidates, which would harm our ability to conduct future clinical trials and, if approved, commercialize our product candidates and generate product revenues in a timely manner or at all.
As we continue to develop and scale our manufacturing processes, we expect that we will need to obtain rights to and supplies of certain materials and equipment to be used as part of those processes. We may not be able to obtain rights to such materials on commercially reasonable terms, or at all, and our inability to alter our processes in a commercially viable manner to avoid the use of such materials or equipment or find suitable substitutes would have a material adverse effect on our business. Even if we are able to alter our processes so as to use other materials or equipment, such a change may delay our clinical development or commercialization plans. If such a change occurs for product candidate that is already being tested in clinical trials, the change may require us to perform comparability studies and to collect additional data from patients prior to undertaking more advanced clinical trials.
We will depend on timely and successful enrollment and retention of patients in our clinical trials for our product candidates. If we experience delays or difficulties enrolling or retaining patients in our clinical trials, our research and development efforts and business, financial condition, and results of operations could be materially adversely affected.
Successful and timely initiation and completion of clinical trials will require that we enroll and retain a sufficient number of patients. Any clinical trials we conduct may be subject to delays for a variety of reasons, including as a result of patient enrollment taking longer than anticipated, patient withdrawal, or the occurrence of adverse events. These types of developments could cause us to delay the trial or halt further development of the relevant product candidate.
Our clinical trials will compete with other clinical trials that are in the same therapeutic areas as our product candidates, and this competition will reduce the number and types of patients available to participate in our trials, as some patients who might have opted to enroll in our trials may instead opt to enroll in a trial being conducted by one of our competitors. Moreover, enrolling patients in clinical trials for diseases in which there is an approved standard of care is challenging, as patients will first receive the applicable standard of care. Many patients who respond positively to the standard of care do not enroll in clinical trials. This may limit the number of eligible patients who have the potential to benefit from our product candidates and could extend development timelines or increase costs for our programs. Patients who fail to respond positively to the standard of care treatment would be eligible for clinical trials of our product candidates. However, treatment with prior regimens may render our product candidates less effective in clinical trials.
Because the number of qualified clinical investigators and clinical trial sites is limited, we expect to conduct at least some of our clinical trials at the same clinical trial sites that some of our competitors use, which will reduce the number of patients who are available for our clinical trials at such clinical trial sites.
Patient enrollment in clinical trials depends on many factors, including:
|
|
• |
the size and nature of the patient population; |
96
|
|
• |
the severity of the disease under investigation; |
|
|
• |
eligibility criteria for the trial; |
|
|
• |
the proximity of patients to clinical sites; |
|
|
• |
the design of the clinical protocol; |
|
|
• |
the ability to obtain and maintain patient consents; |
|
|
• |
perceived risks and benefits of the product candidate under evaluation, including any perceived risks associated with stem cell-derived product candidates; |
|
|
• |
the ability to recruit clinical trial investigators with the appropriate competencies and experience; |
|
|
• |
the risk that enrolled patients will drop out of the trial before administration of our product candidate or trial completion; |
|
|
• |
the availability of competing clinical trials; |
|
|
• |
the availability of patients during the ongoing COVID-19 pandemic; |
|
|
• |
the availability of new drugs approved for the indication the clinical trial is investigating; and |
|
|
• |
clinicians’ and patients’ perceptions as to the potential advantages of the product candidate being studied in relation to other available therapies. |
These factors may make it difficult for us to enroll enough patients to complete our clinical trials in a timely and cost-effective manner. Delays in the completion of any clinical trial of our product candidates will increase our costs, slow down our product candidate development and approval process, and delay or potentially jeopardize our ability to commence product sales and generate revenue. In addition, some of the factors that may cause, or lead to, a delay in the commencement or completion of clinical trials may also ultimately lead to the denial of regulatory approval of our product candidates.
We may become exposed to costly and damaging liability claims, either when testing our product candidates in clinical trials or at the commercial stage, and our product liability insurance may not cover all damages arising from such claims.
We are exposed to potential product liability and professional indemnity risks that are inherent in the research, development, manufacturing, marketing, and use of pharmaceutical products. While we currently have no product candidates for which we have commenced clinical trials or obtained approval for commercial sale, the future use of our product candidates in clinical trials, and the sale of any products for which we may obtain approval in the future, may expose us to liability claims. These claims might be made by patients that use the product, healthcare providers, pharmaceutical companies, or others selling such products. Any claims against us, regardless of their merit, could be difficult and costly to defend and could materially adversely affect the market for our product candidates or any prospects for commercialization of our product candidates.
Although the clinical trial process is designed to identify and assess potential side effects, it is always possible that a drug, even after regulatory approval, may exhibit unforeseen side effects. Physicians and patients may not comply with any warnings that identify known potential adverse effects or patients who should not use our product candidates. If any of our product candidates were to cause adverse side effects during clinical trials or after approval, we may be exposed to substantial liabilities.
We would require significant financial and management resources to defend against any product liability claims, even if we are successful in such defense. Regardless of the merits or eventual outcome, liability claims may result in decreased demand for our product candidates, negative publicity and injury to our reputation, withdrawal of clinical trial participants, initiation of investigations by regulatory authorities, costs to defend the related litigation, diversion of management’s time and our resources, substantial monetary awards to clinical trial participants or patients, product recalls, withdrawals, or labeling, marketing, or promotional restrictions, loss of revenue, exhaustion of any available insurance and our capital resources, inability to commercialize our product candidates, and a decline in our share price.
Although we maintain product liability insurance for our product candidates, it is possible that our liabilities could exceed our insurance coverage. We intend to expand our insurance coverage to include the sale of commercial products if we obtain marketing approval for any of our product candidates. However, we may be unable to maintain insurance coverage at a reasonable cost or obtain insurance coverage that will be adequate to satisfy any liability that may arise. If a successful product liability claim or series of claims is brought against us for uninsured liabilities or in excess of insured liabilities, our assets may not be sufficient to cover such claims, and our business operations could be impaired.
97
Risks Related to Our Dependence on Third Parties
We rely on, and expect to continue to expect to rely on, CDMOs to manufacture our product candidates, as well as materials used in the manufacturing of our product candidates. Any failure by a CDMO to produce acceptable materials or product candidates for us or any failure by us or such manufacturer to obtain authorization from the FDA or comparable foreign regulatory authorities or otherwise satisfy regulatory requirements with respect to such manufacturing of our product candidates may delay or impair our ability to initiate or complete our clinical trials, obtain regulatory approvals, or commercialize approved products.
We do not currently own or operate any cGMP manufacturing facilities, nor do we have any in-house cGMP manufacturing capabilities. In July 2021, we entered into a long-term lease to establish and operate our own cGMP manufacturing facility to support our late-stage clinical development and early commercial activities across our product portfolio. Though we plan to begin building out this facility in the near future, we expect that it will take several years before we are able to begin manufacturing our product candidates at this facility, if at all. Until we are able to begin manufacturing our product candidates at our facility, we will rely on CDMOs to manufacture our product candidates for preclinical and clinical testing and will continue to rely on CDMOs to manufacture certain of our product candidates thereafter as part of our manufacturing strategy. A limited number of CDMOs specialize in or have the expertise required to manufacture our product candidates. Moreover, our CDMOs have limited capacity at their facilities and require commitments to secure availability well in advance of manufacturing any products. Additionally, we face competition from other biopharmaceutical companies to secure availability to manufacture our product candidates at these facilities. If the CDMOs on which we rely to manufacture our product candidates do not have sufficient availability at their facilities to manufacture our product candidates in accordance with our timelines or are not otherwise able to meet our expected deadlines, we will experience delays in manufacturing our product candidates. In addition, our CDMOs face intense competition to attract and retain qualified personnel. If our CDMOs are unable to attract, retain, and motivate qualified personnel, they may be unable to perform their obligations in a timely manner, or their performance may be substandard or may not meet our quality requirements, which could cause us to experience delays in manufacturing our product candidates. Any delays in manufacturing our product candidates could materially harm our ability to conduct our clinical trials or commercialize our product candidates in a timely manner or at all and could harm our business.
In addition, we rely on multiple CDMOs to produce sufficient quantities of materials required for the manufacture of our product candidates for preclinical testing and future clinical trials, and intend to continue to rely on such CDMOs for the commercial manufacture of certain of our products, if approved. If we are unable to arrange for such third-party manufacturing sources, or fail to do so on commercially reasonable terms, we may not be able to produce sufficient supply of product candidate or we may be delayed in doing so. Such failure or substantial delay could materially harm our business.
We rely on third parties for biological materials that are used in our discovery and development programs. These materials can be difficult to produce and occasionally have variability from our product specifications. If these materials do not comply with our product specifications, or in the event of any other disruption in the supply of these materials, our business could be materially adversely affected. Although we have control processes and screening procedures, biological materials are susceptible to damage and contamination and may contain active pathogens. We may also have low yield from certain manufacturing batches, which could increase our costs and slow our development timelines. Improper storage of these materials, by us or any third-party suppliers, may require us to destroy some of these materials or product candidates generated using such materials.
Reliance on CDMOs entails risks to which we would not be subject if we manufactured product candidates ourselves, including reliance on the CDMO for regulatory compliance and quality control and assurance, volume production, the possibility of breach of the manufacturing agreement by the CDMO due to factors beyond our control (including a failure to synthesize and manufacture our product candidates in accordance with our product specifications), and the possibility of termination or nonrenewal of the agreement by the CDMO at a time that is costly or damaging to us.
In addition, the FDA and comparable foreign regulatory authorities require that our product candidates be manufactured according to cGMPs and similar foreign standards relating to methods, facilities, and controls used in the manufacturing, processing, and packing of the product candidate, which are intended to ensure that biological products are safe and that they consistently meet applicable requirements and specifications.
Pharmaceutical manufacturers are required to register their facilities and products manufactured at the time of submission of the marketing application and then annually thereafter with the FDA and certain state and foreign agencies. If the FDA or a comparable foreign regulatory authority does not approve our CDMO’s facilities for the manufacture of our product candidates or if it withdraws any such approval in the future, we may need to find alternative manufacturing facilities, which would significantly impact our ability to develop, obtain regulatory approval for, or market our product candidates, if approved, on a timely basis or at all. Any discovery of problems with a product, or a manufacturing or laboratory facility used by us or our strategic partners in connection with manufacturing of that product, may result in restrictions on the product or on the relevant facility, including marketed product recall, suspension of manufacturing, product seizure, or a voluntary withdrawal of the drug from the market. We may have little to no control regarding the occurrence of any such incidents at our CDMOs.
98
If we were unable to timely find an adequate replacement for our CDMOs or another acceptable solution, our clinical trials could be delayed, or our commercial activities could be harmed. In addition, because we are dependent on our collaborators, our suppliers, and other third parties for the manufacture, filling, storage, and distribution of our product candidates, we have limited ability to prevent or control manufacturing defects in our products. The sale of products containing such defects could adversely affect our business, financial condition, and results of operations. Any failure by our CDMOs to comply with cGMP or failure to properly scale-up manufacturing processes for our product candidates, or any failure to deliver sufficient quantities of product candidates in a timely manner, could lead to a delay in, or failure to obtain, regulatory approval of any of our product candidates.
Pharmaceutical manufacturers are also subject to extensive post-marketing oversight by the FDA and comparable regulatory authorities in the jurisdictions where a product is marketed, which includes periodic unannounced and announced inspections by the FDA to assess compliance with cGMP requirements. Any failure by one of our CDMOs to comply with cGMP or to provide adequate and timely corrective actions in response to deficiencies identified in a regulatory inspection could result in further enforcement action that could lead to a shortage of products and harm our business, including withdrawal of approvals previously granted, seizure, injunction, or other civil or criminal penalties. The failure of a CDMO to address any concerns raised by the FDA or comparable foreign regulatory authorities could also lead to plant shutdown or the delay or withholding of product approval by the FDA in additional indications or by comparable foreign regulatory authorities in any indication. Certain countries may impose additional requirements on the manufacturing of drug products or drug substances, and on manufacturers, as part of the regulatory approval process for products in such countries. The failure by our CDMOs to satisfy such requirements could impact our ability to obtain or maintain approval of our products in such countries.
If we are unable to obtain sufficient raw and intermediate materials on a timely basis or if we experience other manufacturing or supply interruptions or difficulties, we may be unable to resume supply of such materials or other manufacturing activities within a reasonable time frame and at an acceptable cost or at all, which would adversely affect our business.
The manufacture of our product candidates requires the timely delivery of sufficient amounts of raw and intermediate materials. We purchase, and rely on our CDMOs to purchase, certain of these materials from third-party suppliers in order to produce our product candidates for our preclinical studies. There are a limited number of suppliers of these materials, and we may need to assess alternate suppliers to prevent possible disruption of manufacturing of our product candidates for our preclinical studies, our future clinical trials, and if ultimately approved, commercial sale. We intend to continue to rely on our CDMOs to purchase materials in order to produce product candidates for any clinical trials that we undertake; however, we do not have any control over the process or timing of the acquisition of these materials by our CDMOs. We work closely with our CDMOs and suppliers, as applicable, to ensure the continuity of supply, but cannot ensure that these efforts will always be successful. Further, while we strive to diversify our sources of raw and intermediate materials, in certain instances we acquire raw and intermediate materials from a sole supplier. We cannot be sure that these suppliers will remain in business, or that they will not be purchased by one of our competitors or another company that is not interested in continuing to supply these materials for our intended purpose. In addition, the lead time needed to establish a relationship with a new supplier can be lengthy, and we may experience delays in the event a new supplier must be used. The time and effort to qualify a new supplier could result in additional costs, diversion of resources, or reduced manufacturing yields, any of which would negatively impact our operating results. Although we generally would not begin a clinical trial unless we believe we have a sufficient supply of a product candidate to complete the clinical trial, any significant delay in the supply of a product candidate, or the raw or intermediate material components thereof, for an ongoing clinical trial due to the need to replace a third-party manufacturer could considerably delay completion of our clinical trials, product testing, and potential regulatory approval of our product candidates. While we believe that alternative sources of supply exist where we rely on sole supplier relationships, we cannot ensure that, if needed, we would be able to quickly establish additional or replacement sources for some materials. Moreover, we currently do not have any agreements for the commercial production of these raw or intermediate materials. If any of our product candidates receives regulatory approval and thereafter, we or our CDMOs are unable to purchase these raw or intermediate materials, the commercial launch of our product candidates could be delayed or there could be a shortage in supply of product, which would impair our ability to generate revenues from the sale of such approved product. A reduction or interruption in supply of raw or intermediate materials, and an inability to establish alternative sources for such supply, could adversely affect our ability to manufacture our product candidates or approved products in a timely or cost-effective manner.
We rely, and expect to continue to rely, on third parties, including independent clinical investigators and CROs, to conduct or support our preclinical studies and clinical trials. If these third parties do not successfully carry out their contractual duties, comply with applicable regulatory requirements, or meet expected deadlines, we may not be able to obtain regulatory approval for or commercialize our product candidates and our business could be substantially harmed.
We have relied upon and plan to continue to rely upon third parties, including independent clinical investigators and third-party CROs, to conduct or support our preclinical studies and clinical trials and to monitor and manage data for our ongoing preclinical and clinical programs. We rely or will rely on these parties for execution of our preclinical studies and clinical trials and control only certain aspects of their activities. Even then, we are only able to control such activities to the extent set forth under our contracts with the relevant third parties. Nevertheless, we are responsible for ensuring that each of our preclinical and clinical studies and trials is
99
conducted in accordance with the applicable protocol and legal, regulatory, and scientific standards and rules, and our reliance on these third parties does not relieve us of these obligations. With respect to any of our product candidates that may enter clinical development, we and our third-party contractors and CROs are required to comply with GCP requirements, which are regulations and guidelines enforced by the FDA and comparable foreign regulatory authorities. Regulatory authorities enforce these GCPs through periodic inspections of clinical trial sponsors, principal investigators, and clinical trial sites. If we or any of our CROs, or any principal investigators or clinical trial sites involved in our trials, fail to comply with applicable GCPs, the clinical data generated from these clinical trials may be deemed unreliable and the FDA or comparable foreign regulatory authorities may require us to perform additional clinical trials before approving our marketing applications. We cannot be certain that upon inspection by a given regulatory authority, such regulatory authority will determine that any of our clinical trials comply with GCP regulations. In addition, our clinical trials must be conducted with product produced under cGMP regulations. Our failure to comply with these regulations may require us to repeat clinical trials, which would delay the regulatory approval process for the relevant product candidate.
Further, principal investigators, clinical trial sites and CROs are not our employees, and we are unable to control, other than by contract, the amount of resources, including time, that they devote to our product candidates and clinical trials. If our CROs are unable to attract, retain, and motivate qualified personnel, they may be unable to perform their obligations in a timely manner, or their performance may be substandard. If principal investigators, clinical trial sites or CROs fail to devote sufficient resources to the development of our product candidates, or if their performance is substandard or does not meet our quality requirements, it may delay or compromise the prospects for approval and commercialization of any such product candidates. In addition, in order for these third parties to perform under their contracts with us, we regularly disclose or plan to disclose to these third parties confidential or proprietary information, which increases the risk that this information will be misappropriated. Additionally, disruptions caused by global geo-political, economic, and other factors beyond our control, including the ongoing COVID-19 pandemic, may increase the likelihood that these third parties encounter difficulties or delays in performing their obligations to us, including with respect to initiating, enrolling, conducting, or completing our planned clinical trials. In particular, we have experienced and may continue to experience difficulty in accessing animal models, specifically non-human primate models, for the preclinical evaluation of our product candidates. Delays caused by the inability to access these models may cause our development timelines to be extended beyond what we anticipate.
Third parties, including our CROs, generally have the right to terminate their agreements with us in the event of an uncured material breach by us. In addition, certain third parties may have the right to terminate their respective agreements with us under other circumstances, including if it can be reasonably demonstrated that the safety of the subjects participating in our clinical trials warrants such termination, if we make a general assignment for the benefit of our creditors, or if we are liquidated.
There is a limited number of third parties, including service providers and clinical trial sites, that specialize or have the expertise required to achieve our business objectives. If any of our relationships with these third-parties, including laboratories, CROs, or clinical trial sites, terminate, we may not be able to enter into arrangements with alternative third parties or to do so in a timely manner or on commercially reasonable terms. If these third parties do not successfully carry out their contractual duties or obligations or meet expected deadlines, if they need to be replaced, or if the quality or accuracy of the data they obtain is compromised due to the failure to adhere to our preclinical or clinical protocols, regulatory requirements, or for other reasons, our preclinical or clinical trials may be extended, delayed, or terminated and we may not be able to obtain regulatory approval for or successfully commercialize our product candidates. As a result, our results of operations and the commercial prospects for our product candidates would be harmed, our costs could increase, and our ability to generate revenues could be delayed. Switching from existing service providers or clinical trial sites, or adding additional service providers or clinical trial sites, involves additional cost and requires management time and focus. In addition, there is a natural transition period when a new service provider commences work. As a result, delays can occur, which may materially impact our ability to meet our desired development, including clinical development, timelines. Additionally, service providers may lack the capacity to absorb higher workloads or take on additional capacity to support our needs. Though we carefully manage our relationships with these service providers, including our contracted laboratories and CROs, there can be no assurance that we will not encounter these types of challenges or delays in the future or that these delays or challenges will not have a material adverse impact on our business, financial condition, and prospects.
In addition, clinical investigators may serve as scientific advisors or consultants to us from time to time and may receive cash or equity compensation in connection with such services. If these relationships and any related compensation result in perceived or actual conflicts of interest, or the FDA concludes that the financial relationship may have affected the conduct or interpretation of one of our preclinical studies or clinical trials, the integrity of the data generated from such preclinical study or clinical trial may be questioned and the utility of the preclinical study or clinical trial itself may be jeopardized, which could result in the delay or rejection by the FDA of any regulatory submissions related to our product candidates. Any such delay or rejection could prevent us from commercializing our product candidates.
100
Risks Related to Intellectual Property and Information Technology
We may not be able to protect our intellectual property rights throughout the world.
Patent rights are national or regional rights. The filing, prosecution, maintenance, and defense of patent rights on our platform technologies and product candidates worldwide would be prohibitively expensive, and our intellectual property rights in some countries outside the United States may have a different scope and strength than do those in the United States. In addition, the laws of some foreign countries, particularly certain developing countries, do not protect intellectual property rights to the same extent as federal and state laws in the United States. Consequently, we may not be able to prevent third parties from practicing our intellectual property rights in all countries outside the United States or from making, using, selling, or importing products made using our intellectual property rights in and into the United States or other jurisdictions. Competitors may use our technologies in jurisdictions where we have not obtained intellectual property rights, including patent protection, to develop their own products and may also export otherwise infringing products to territories where we have intellectual property rights, including patent protection, but enforcement rights are not as strong as those in the United States. These products may compete with our products and our patent or other intellectual property rights may not be effective or adequate to prevent them from competing.
Many companies have encountered significant problems in protecting and defending intellectual property rights in foreign jurisdictions. The legal systems of certain countries, particularly certain developing countries, do not favor the enforcement of patents, trade secrets, and other intellectual property, particularly those relating to biopharmaceutical products, which could make it difficult in those jurisdictions for us to stop the infringement or misappropriation of our patents or other intellectual property rights, or the marketing of competing products in violation of our proprietary rights. Proceedings to enforce our patent and other intellectual property rights in foreign jurisdictions are expensive, especially in jurisdictions where we have no local presence, and could result in substantial costs and divert our efforts and attention from other aspects of our business. Furthermore, such proceedings could put our patents at risk of being invalidated, held unenforceable, or interpreted narrowly, could put our patent applications at risk of not issuing, and could provoke third parties to assert claims of infringement or misappropriation against us. We may not prevail in any lawsuits that we initiate, and the damages or other remedies awarded, if any, may not be commercially meaningful. Similarly, if our trade secrets are disclosed in a foreign jurisdiction, competitors worldwide could have access to our proprietary information, and we may be without satisfactory recourse. Such disclosure could have a material adverse effect on our business. Moreover, our ability to protect and enforce our intellectual property rights may be adversely affected by unforeseen changes in foreign intellectual property laws. In addition, certain developing countries, including China and India, have compulsory licensing laws under which a patent owner may be compelled to grant licenses to third parties. In those countries, we and our licensors may have limited remedies if patents are infringed or if we or our licensors are compelled to grant a license to a third-party, which could materially diminish the value of those patents. In addition, many countries limit the enforceability of patents against government agencies or government contractors. This could limit our potential revenue opportunities. Accordingly, our efforts to enforce our intellectual property rights around the world may be inadequate to obtain a significant commercial advantage from the intellectual property that we develop or license.
Because of the expense and uncertainty of litigation, we may conclude that, even if a third party is infringing our issued patents, or any patents that may be issued as a result of our pending or future patent applications, or other intellectual property rights, the risk-adjusted cost of bringing and enforcing such a claim or action, which typically lasts for years before it is concluded, may be too high or not in the best interest of our company or our stockholders, or it may be otherwise impractical or undesirable to enforce our intellectual property against some third parties. Our competitors or other third parties may be able to sustain the costs of complex patent litigation or proceedings more effectively than we can because of their greater financial resources and/or more mature and developed intellectual property portfolios. In such cases, we may decide that the monetary cost of such litigation and the diversion of the attention of our management and scientific personnel could outweigh any benefit we may receive as a result of the proceedings and that the more prudent course of action is to simply monitor the situation or initiate or seek some other non-litigious action or solution. In addition, the uncertainties associated with litigation could compromise our ability to raise the funds necessary to initiate or continue our future clinical trials, continue our internal research programs, in-license needed technology or other product candidates, or enter into development partnerships that would help us bring our product candidates to market.
We depend on intellectual property licensed from third parties, and our rights to develop and commercialize our product candidates are subject to, in part, the terms and conditions of the licenses granted to us by such third parties. If we breach our obligations under these agreements or if any of these agreements is terminated, we may be required to pay damages, lose our rights to such intellectual property and technology, or both, which would harm our business.
We depend on patents, know-how, and proprietary technology, both our own and those that we license from others to research, develop, and commercialize our product candidates. We are a party to a number of intellectual property license agreements and acquisition agreements pursuant to which we have acquired certain of our core intellectual property rights. Moreover, we rely upon licenses to certain intellectual property rights and proprietary technology from third parties that are important or necessary for the development of our technology and products, including technology related to our manufacturing processes and our product candidates. These licenses may not provide exclusive rights to use such intellectual property and technology in all relevant fields of use or in all territories in which we may wish to develop or commercialize our technology and products in the future. As a result, we may not be
101
able to prevent competitors from developing and commercializing competitive products in such fields of use or territories. These licenses may also require us to grant back certain intellectual property rights to our licensors and to pay certain amounts relating to sublicensing patent and other rights.
In the future, we expect to enter into additional license agreements. For example, with respect to our ex vivo cell engineering platform relying on hypoimmune technology, we have licensed certain intellectual property from Harvard, UCSF, and Washington University. Additionally, we acquired our in vivo cell engineering platform, which is based on fusogen technology, from Cobalt, which included several license agreements and options-to-license, as well as our glial progenitor cell and cardiomyocyte programs from Oscine and Cytocardia, respectively, both of which came with in-licenses. These license and acquisition agreements impose, and we expect that future license and acquisition agreements will impose, various diligence, milestone, and royalty payment, and other obligations on us. If we fail to comply with our obligations under these agreements, we may be required to pay damages, and the licensor may have the right to terminate the agreement. Any termination of these licenses could result in the loss of significant rights and could harm our ability to develop or advance one of our cell engineering platforms, or develop, manufacture, or commercialize one of our product candidates. See the subsection titled “Business— Key Intellectual Property Agreements” in Part I, Item 1 of this Annual Report for additional information regarding these key agreements.
In addition, the agreements under which we license intellectual property or technology to or from third parties are complex, and certain provisions in such agreements may be susceptible to multiple interpretations. The resolution of any contract interpretation disagreement that may arise could narrow what we believe to be the scope of our rights to the relevant intellectual property or technology or increase what we believe to be our financial or other obligations under the relevant agreement, either of which could have a material adverse effect on our business, financial condition, results of operations, and prospects. Moreover, if disputes over intellectual property that we have licensed prevent or impair our ability to maintain our current licensing arrangements on commercially acceptable terms, we may be unable to successfully develop and commercialize the affected product candidates. Our business would also suffer if any current or future licensors fail to abide by the terms of the license, if such licensors fail to enforce licensed patents against infringing third parties, if the licensed patents or other rights are found to be invalid or unenforceable, or if we are unable to enter into necessary licenses on acceptable terms or at all. Moreover, our licensors may own or control intellectual property that has not been licensed to us and, as a result, we may be subject to claims, regardless of their merit, that we are infringing or otherwise violating the licensor’s rights.
In addition, while we cannot currently determine the amount of the royalty obligations we would be required to pay on sales of future products, if any, the amounts may be significant. The amount of our future royalty obligations will depend on the technology and intellectual property we use in product candidates that we successfully develop and commercialize, if any. Therefore, even if we successfully develop and commercialize any product candidates, we may be unable to achieve or maintain profitability.
If we are unable to successfully maintain the existing intellectual property rights we have, we may have to abandon development of the relevant research programs or product candidates, and our business, financial condition, results of operations and prospects could suffer.
Licensing of intellectual property is of critical importance to our business, involves complex legal, business, and scientific issues and is complicated by the rapid pace of scientific discovery in our industry. Disputes may also arise between us and our licensors regarding intellectual property subject to a license agreement, including those relating to:
|
|
• |
the scope of rights granted under the license agreement and other interpretation-related issues; |
|
|
• |
whether and the extent to which our technology and processes infringe on intellectual property of the licensor that is not subject to the license agreement; |
|
|
• |
our right to sublicense the patent and other rights granted to us under the license agreement to third parties as part of collaborative development relationships; |
|
|
• |
whether we are complying with our diligence obligations with respect to the use of the licensed intellectual property rights in relation to our development and commercialization of our product candidates, and what activities satisfy those diligence obligations; |
|
|
• |
the priority of invention of patented technology; |
|
|
• |
the amount and timing of payments owed under license agreements; and |
|
|
• |
the allocation of ownership of inventions and know-how resulting from the joint creation or use of intellectual property by our licensors and by us and our partners. |
If disputes over intellectual property that we have licensed prevent or impair our ability to maintain our current licensing arrangements on acceptable terms, we may be unable to successfully develop and commercialize the affected product candidates. We
102
are generally also subject to all of the same risks with respect to protection of intellectual property that we license as we are for intellectual property that we own, which are described below. If we or our licensors fail to adequately protect this intellectual property, our ability to commercialize our products could suffer.
We depend, in part, on our licensors to file, prosecute, maintain, defend, and enforce certain patents and patent applications that are material to our business.
Certain patents relating to our product candidates are owned or controlled by certain of our licensors. Each of our licensors generally has rights to file, prosecute, maintain, and defend the patents we have licensed from such licensor in their name, generally with our right to comment on such filing, prosecution, maintenance, and defense, with some obligation for the licensor to consider or incorporate our comments, for our exclusively licensed patents. We generally have the first right to enforce our exclusively licensed patent rights against third parties, although our ability to settle such claims often requires the consent of the licensor. If our licensors, third parties from whom they license or have obtained the relevant patents, or any future licensees having rights to file, prosecute, maintain, and defend our patent rights fail, or have in the past failed, to properly and timely conduct these activities for patents or patent applications covering any of our product candidates, including due to the impact of the COVID-19 pandemic on our licensors’ or such third parties’ business operations, our ability to develop and commercialize those product candidates may be adversely affected and we may not be able to prevent competitors from making, using, or selling competing products. We cannot be certain that such activities have been or will be conducted in compliance with applicable laws and regulations or will result in valid and enforceable patents or other intellectual property rights. Pursuant to the terms of the license agreements with some of our licensors, these licensors may have the right to control enforcement of our licensed patents or defense of any claims asserting the invalidity of these patents and, even if we are permitted to pursue such enforcement or defense, we cannot ensure the cooperation of our licensors. We cannot be certain that our licensors will allocate sufficient resources or prioritize their or our enforcement of such patents or defense of such claims to protect our interests in the licensed patents. Even if we are not a party to these legal actions, an adverse outcome could harm our business because it could cause us to lose rights to intellectual property that we may need to operate our business or could cause us to lose the ability to exclude our competitors from using the intellectual property rights. In addition, even when we have the right to control patent prosecution of licensed patents and patent applications, enforcement of licensed patents, or defense of claims asserting the invalidity of those patents, we may still be adversely affected or prejudiced by actions or inactions of our licensors and their counsel that took place prior to or after our assuming control. In the event we breach any of our contractual obligations to our licensors related to such prosecution, we may incur significant liability to our licensors.
We may not be successful in obtaining or maintaining necessary rights to product candidates, product candidate components, or processes for our product development pipeline, which may require us to operate our business in a more costly or otherwise adverse manner than we anticipated. We may not be successful in obtaining or maintaining exclusive rights to owned and in-licensed patents or patent applications or future patents to the extent they are co-owned with third parties.
We own or license from third parties certain intellectual property rights necessary to develop our product candidates. The growth of our business will likely depend in part on our ability to acquire or in-license additional proprietary rights, including to advance our research or allow commercialization of our product candidates. If we are unable to do so, we may be required to expend considerable time and resources to develop or license replacement technology. For example, our programs may rely upon additional technologies or product candidates that require the use of additional proprietary rights held by third parties. Furthermore, other pharmaceutical companies and academic institutions may have filed or may plan to file patent applications potentially relevant to our business. In order to work effectively and efficiently, our product candidates may also require specific formulations or other technology, which may be covered by intellectual property rights held by others. From time to time, in order to avoid infringing third-party patents, we may be required to license technology from these third parties to further develop or commercialize our product candidates. We may be unable to acquire or in-license third-party intellectual property rights that we identify as necessary or important to our business operations, including those required to make, use, or sell our product candidates. We may fail to obtain any of these licenses at a reasonable cost or on reasonable terms, if at all, and, as a result, we may be unable to develop or commercialize the affected product candidates, which would harm our business. We may need to cease use of the compositions or methods covered by such third-party intellectual property rights. In addition, may need to seek to develop alternative approaches that do not infringe on such intellectual property rights, which, if we were successful in developing such alternatives, may entail additional costs and lead to delays in development. In certain cases, it may not be feasible for us to develop such alternatives. Even if we are able to obtain a license under such intellectual property rights, any such license may be non-exclusive, which may allow our competitors to access to the same technologies licensed to us.
Additionally, we sometimes collaborate with academic institutions to accelerate our preclinical research or development under written agreements with these institutions. Typically, these institutions provide us with an option to negotiate a license to any of the institution’s rights in technology resulting from the collaboration. Regardless of such option, we may be unable to negotiate a license within the specified timeframe or under terms that are acceptable to us. If we are unable to do so, the institution may license the intellectual property rights to other parties, potentially blocking our ability to pursue any of our programs to which such rights relate. If we are unable to successfully obtain rights to required third-party intellectual property or to maintain the existing intellectual
103
property rights we have, we may have to abandon development of one or more programs and our business and financial condition could suffer.
The licensing and acquisition of third-party intellectual property rights is competitive, and companies that may be more established, or have greater resources than we do, may also be pursuing strategies to license or acquire third-party intellectual property rights that we may consider necessary or attractive in order to commercialize our product candidates. More established companies may have a competitive advantage over us due to their larger size and cash resources or greater clinical development and commercialization capabilities. There can be no assurance that we will be able to successfully complete negotiations and ultimately license or acquire the rights to the intellectual property necessary or useful for the development of our product candidates.
Our product candidates may also require specific components to work effectively and efficiently, and rights to those components may be held by third parties. We may be unable to in-license any compositions, methods of use, processes, or other intellectual property rights from any such third parties that we identify, including because such licenses may not be available at a reasonable cost or on reasonable terms, which would harm our business. Even if we are able to obtain a license, it may be non-exclusive, thereby giving our competitors access to the same technologies licensed to us. In that event, we may be required to expend significant time and resources to develop or license replacement technology. Any delays in entering into new collaborations or strategic partnership agreements related to our product candidates could delay the development and commercialization of our product candidates in certain geographies, which could harm our business prospects, financial condition, and results of operations.
Moreover, some of our owned and in-licensed patents or patent applications or future patents are or may be co-owned with third parties. If we are unable to obtain an exclusive license to any such third-party co-owner’s interest in such patents or patent applications, such co-owner may be able to license their rights to other third parties, including our competitors, and our competitors could market competing products and technologies. In addition, we may need the cooperation of any such co-owner in order to enforce such patents against third parties, and such cooperation may not be provided to us. Furthermore, our owned and in-licensed patents may be subject to a reservation of rights by one or more third parties. Any of the foregoing could have a material adverse effect on our competitive position, business, financial conditions, results of operations, and prospects.
We may depend on intellectual property licensed or sublicensed to us from, or for which development was funded or otherwise assisted by, government agencies, such as the National Institutes of Health, for development of our technology and product candidates.
Government agencies have provided and may in the future provide funding, facilities, personnel, or other assistance in connection with the development of the intellectual property rights owned by or licensed to us. Such government agencies may have retained rights in such intellectual property, including the right to grant or require us to grant mandatory licenses or sublicenses to such intellectual property to third parties under certain specified circumstances, including if it is necessary to meet health and safety needs that we are not reasonably satisfying or if it is necessary to meet requirements for public use specified by federal regulations, or to manufacture products in the United States. Any exercise of such rights, including with respect to any such required sublicense of these licenses, could result in the loss of significant rights and could harm our ability to commercialize or continue commercializing licensed products. For example, at least one of our in-licensed patent cases related to each of our ex vivo cell engineering and in vivo cell engineering platforms has been funded at least in part by the United States government. As a result, these patent cases are subject to certain federal regulations pursuant to the Bayh-Dole Act of 1980 (Bayh-Dole Act). In particular, the federal government retains a “nonexclusive, nontransferable, irrevocable, paid-up license” for its own benefit to inventions produced with its financial assistance. The Bayh-Dole Act also provides federal agencies with “march-in rights.” March-in rights allow the government, in specified circumstances, to require the contractors or successors in title to the patent to grant a “nonexclusive, partially exclusive, or exclusive license” to a “responsible applicant or applicants.” If the patent owner refuses to do so, the government may grant the license itself. Intellectual property rights discovered under government-funded programs are also subject to certain reporting requirements, compliance with which may require us or our licensors to expend substantial resources and failure to comply may lead to loss of rights. Such intellectual property is also subject to a preference for United States industry, which may limit our ability to contract with foreign product manufacturers for products covered by such intellectual property rights. Moreover, we sometimes collaborate with academic institutions to accelerate our preclinical research or development, and we cannot be sure that any co-developed intellectual property will be free from government rights pursuant to the Bayh-Dole Act. If, in the future, we co-own or in-license technology that is critical to our business and is developed in whole or in part with federal funds subject to the Bayh-Dole Act, our ability to enforce or otherwise exploit patents covering such technology may be adversely affected.
104
If we are unable to obtain and maintain sufficient intellectual property protection for our platform technologies and product candidates, or if the scope of the intellectual property protection is not sufficiently broad, our competitors could develop and commercialize products similar or identical to ours, and our ability to successfully commercialize our products may be adversely affected.
We anticipate that we will file additional patent applications both in the United States and in other countries, as appropriate. However, we cannot predict:
|
|
• |
if and when any patents will issue; |
|
|
• |
the degree and range of protection any issued patents will afford us against competitors, including whether third parties will find ways to invalidate or otherwise circumvent our patents; |
|
|
• |
whether others will apply for or obtain patents claiming inventions similar to those covered by our patents and patent applications; |
|
|
• |
whether we will need to initiate litigation or administrative proceedings to defend our patent rights, which may be costly whether we win or lose; or |
|
|
• |
whether the patent applications that we own or in-license will result in issued patents with claims that cover our product candidates or uses thereof in the United States or in other foreign countries. |
Our success depends in large part on our ability to obtain and maintain patent protection in the United States and other countries with respect to our platform technologies and product candidates. We seek to protect our proprietary position by filing patent applications in the United States and abroad related to our novel discoveries and technologies that are important to our business.
Obtaining and enforcing patents is expensive and time-consuming, and we may not be able to file, and we and our licensors may not be able to prosecute, all necessary or desirable patent applications or maintain, defend, or enforce patents that may issue based on our patent applications at a reasonable cost or in a timely manner, including as a result of the COVID-19 pandemic impacting our or our licensors’ operations. It is also possible that we will fail to identify patentable aspects of our research and development results before it is too late to obtain patent protection or before another party files a patent application covering the relevant inventions. Although we enter into non-disclosure and confidentiality agreements with parties that have access to patentable aspects of our research and development output, such as our employees, corporate collaborators, outside scientific collaborators, contract research organizations, contract manufacturers, consultants, advisors, and other third parties, any of these parties may breach these agreements and disclose such results before a patent application is filed, thereby jeopardizing our ability to seek patent protection.
Composition of matter patents for biological products such as ex vivo and in vivo cell engineering product candidates often provide a strong form of intellectual property protection for those types of products, as such patents provide protection without regard to any method of use. We cannot be certain, however, that the claims in our pending patent applications covering the composition of matter of our product candidates will be considered patentable by the United States Patent and Trademark Office (USPTO), or by patent offices in foreign countries, or that the claims in any of our issued patents will be considered valid and enforceable by courts in the United States or foreign countries. Method of use patents protect the use of a product for the specified method. This type of patent does not prevent a competitor from making and marketing a product that is identical to our product for an indication that is outside the scope of the patented method. Moreover, even if competitors do not actively promote their products for our targeted indications, physicians may prescribe these products “off-label” for those uses that are covered by our method of use patents. Although off-label prescriptions may infringe or contribute to the infringement of method of use patents, the practice is common and such infringement may be difficult to prevent or prosecute.
One aspect of the determination of patentability of inventions depends on the scope and content of the “prior art,” which is information that was or is deemed available to a person of skill in the relevant art prior to the priority date of the claimed invention. There may be prior art of which we are not aware that may affect the patentability of our inventions or, if issued, affect the validity or enforceability of a patent claim. Further, we may not be aware of all third-party intellectual property rights potentially relating to our product candidates or their intended uses, and as a result, the impact of such third-party intellectual property rights on the patentability of our own patents and patent applications, as well as upon our freedom to operate, is highly uncertain. Because patent applications in the United States and most other countries are typically confidential for a period of 18 months after filing, or may not be published at all, we cannot be certain that we were the first to file any patent application related to our product candidates. As a result, the issuance, scope, validity, enforceability, and commercial value of our patent rights are highly uncertain. Furthermore, for United States patent applications in which all claims are entitled to a priority date before March 16, 2013, an interference proceeding can be provoked by a third party or instituted by the USPTO to determine who was the first to invent any of the subject matter covered by the patent claims of our applications. For United States patent applications containing a claim not entitled to priority before March 16, 2013, there is a greater level of uncertainty in the patent law in view of the passage of the Leahy-Smith America Invents Act (the Leahy-Smith Act),
105
which introduced significant changes to the United States patent laws, including new procedures for challenging pending patent applications and issued patents.
The strength of patents in the biotechnology and pharmaceutical field can be uncertain and evaluating the scope and validity of such patents involves complex legal, factual, and scientific analyses, which may vary based on the jurisdiction in which the analyses are performed. Patents have in recent years been the subject of much litigation in the United States and worldwide, resulting in court decisions, including United States Supreme Court decisions, that have increased uncertainties as to the patentability of certain inventions as well as the enforceability of patent rights in the future. The patent applications that we own or in-license may fail to result in issued patents with claims that cover our platform technologies or our product candidates or uses thereof in the United States or in other foreign countries. Even if the patents do successfully issue, third parties may challenge the patentability, validity, enforceability, or scope thereof, which may result in such patents being narrowed, invalidated, revoked, or held unenforceable. In the event of litigation or administrative proceedings, we cannot be certain that the claims in any of our issued patents will be considered patentable by administrative bodies or valid by courts in either the United States or foreign countries. Furthermore, even if they are unchallenged, our patents and patent applications may not adequately protect our intellectual property or prevent others from designing their products to avoid being covered by our claims. If the breadth or strength of protection provided by the patent filings we hold with respect to our platform technologies or our product candidates is threatened, this could dissuade companies from collaborating with us to develop, and could threaten our ability to successfully commercialize, our product candidates. In addition, the laws of foreign countries may not protect our rights to the same extent as the laws of the United States, or vice versa. Further, as patent rights are time limited, if we encounter delays in our clinical trials, the period of time during which we could market our product candidates under patent protection would be reduced.
We may not identify relevant third-party patents or may incorrectly interpret the relevance, scope, or expiration of a third-party patent, which might adversely affect our ability to develop and market our products.
We cannot guarantee that any of our patent searches or analyses, including the identification of relevant patents, the scope or validity of patent claims, or the expiration of relevant patents, are complete or thorough, nor can we be certain that we have identified each and every third-party patent and pending application worldwide, including in the United States, that is relevant to or necessary for the commercialization of our product candidates in any jurisdiction. The scope of a patent’s claims is determined by an interpretation of the laws of the country in which the patent has been granted, the written disclosure in the patent, and the patent’s prosecution history. Our interpretation of the relevance or the scope of a patent or a pending application may be incorrect, which may negatively impact our ability to market our products. We may incorrectly determine that our products are not covered by a third-party patent or may incorrectly predict whether a third party’s pending application will issue with claims of relevant scope. Our determination of the expiration date of any patent worldwide, including in the United States, that we consider relevant may be incorrect, which may negatively impact our ability to develop and market our product candidates. Our failure to identify and correctly interpret relevant patents may negatively impact our ability to develop and market our products.
Intellectual property rights do not necessarily protect us from all potential threats to our competitive advantage.
The degree of future protection afforded by our intellectual property rights is uncertain because intellectual property rights have limitations and may not adequately protect our business or permit us to maintain our competitive advantage. For example:
|
|
• |
others may be able to make product candidates that are similar to ours but that are not covered by the claims of the patents that we own or have exclusively licensed; |
|
|
• |
we or our licensors or future collaborators might not have been the first to make the inventions covered by certain issued patents or pending patent applications that we own or have exclusively licensed; |
|
|
• |
we or our licensors or future collaborators might not have been the first to file patent applications covering certain of our inventions; |
|
|
• |
others may independently develop similar or alternative technologies or duplicate any of our technologies without infringing our intellectual property rights; |
|
|
• |
our pending patent applications may not lead to issued patents; |
|
|
• |
issued patents that we own or have exclusively licensed may be revoked or may be held invalid, unpatentable, or unenforceable, including as a result of legal challenges; |
|
|
• |
our competitors might conduct research and development activities in countries where we do not have patent rights and then use the information learned from such activities to develop competitive products for sale in our major commercial markets; |
|
|
• |
we may not develop additional proprietary technologies that are patentable; |
106
|
|
• |
we cannot predict the scope of protection of any patent that may issue based on our patent applications, including whether the patent applications that we own or in-license will result in issued patents with claims that cover our product candidates or uses thereof in the United States or in other foreign countries; |
|
|
• |
the claims of any patent that may issue based on our patent applications may not provide protection against competitors or any competitive advantages, or may be challenged by third parties; |
|
|
• |
if we seek to enforce our patents, a court may not hold that our patents are valid, enforceable, or infringed; |
|
|
• |
we may need to initiate litigation or administrative proceedings to enforce or defend our patent rights, which will be costly regardless of outcome; |
|
|
• |
we may choose not to file a patent in order to maintain certain rights as trade secrets or know-how, and a third party may subsequently file a patent covering such intellectual property; |
|
|
• |
we may fail to adequately protect and police our trademarks and trade secrets; and |
|
|
• |
the patent rights of others may have an adverse effect on our business, including if others obtain patents claiming subject matter similar to or improving that covered by our patents and patent applications. |
Should any of these events occur, they could significantly harm our business, results of operations, and prospects.
Confidentiality agreements with employees and third parties may not prevent unauthorized disclosure of trade secrets and other proprietary information.
In addition to the protection afforded by patents, we rely on trade secret protection and confidentiality agreements to protect proprietary know-how that is not patentable or that we elect not to patent, subject matter for which patents are difficult to enforce, and other elements of our product candidates, technology, and product discovery and development processes that involve proprietary know-how, information, or technology that we do not cover through patent protection. Any disclosure, either intentional or unintentional, by our employees, contractors, collaborators, or those of third parties, including those with whom we share our facilities and consultants and vendors that we engage to perform research, clinical trials, or manufacturing activities, or misappropriation by third parties (such as through a cybersecurity breach) of our trade secrets or proprietary or confidential information could enable competitors to duplicate or surpass our technological achievements, thus eroding our competitive position in our market. Because we expect to rely on third parties in the development and manufacture of our product candidates, we must, at times, share trade secrets with them, which increases the possibility that a competitor will discover them or that our trade secrets will be misappropriated or disclosed.
Trade secrets and confidential information, however, can be difficult to protect. We seek to protect our trade secrets, know-how, and confidential information, including our proprietary processes, in part, by entering into confidentiality agreements with our employees, consultants, outside scientific advisors, contractors, and collaborators. We require our employees to enter into written employment agreements containing provisions of confidentiality and obligations to assign to us any inventions generated in the course of their employment. In addition, we enter into agreements with our consultants, contractors, and outside scientific collaborators that typically include invention assignment obligations. We cannot guarantee that we have entered into such agreements with each party that may have or has had access to our trade secrets or proprietary or confidential information, including our technology and processes. Although we use reasonable efforts to protect our trade secrets and confidential information, our employees, consultants, outside scientific advisors, contractors, and collaborators might intentionally or inadvertently disclose such information to competitors, including, as to consultants and advisors, to their primary employers, in breach of our agreements with such parties, and adequate remedies for such breaches may be unavailable. In addition, competitors may otherwise gain access to our trade secrets or independently develop substantially equivalent information and techniques. Enforcing a claim that a party illegally disclosed or misappropriated a trade secret is difficult, expensive, and time-consuming, and the outcome is unpredictable. If any of our trade secrets were to be lawfully obtained or independently developed by a competitor or other third party, we would have no right to prevent them from using that technology or information to compete with us. Furthermore, the laws of some foreign countries do not protect proprietary rights to the same extent or in the same manner as the laws of the United States. As a result, we may encounter significant problems in protecting and defending our intellectual property both in the United States and abroad. If we are unable to prevent unauthorized material disclosure of our intellectual property to third parties or misappropriation of our intellectual property by third parties, we will not be able to establish or maintain a competitive advantage in our market, which could materially adversely affect our business, operating results, and financial condition.
107
Third-party claims of intellectual property infringement against us or our collaborators may prevent or delay our product discovery, development, or commercialization efforts.
Our commercial success depends in part on our avoiding infringement of the patents and proprietary rights of third parties. We cannot be certain that our platform technologies, product candidates, and other proprietary technologies we may develop will not infringe existing or future patents owned by third parties. The legal and administrative landscape related to infringement of the patents and proprietary rights of third parties is fluid as there is a substantial amount of litigation involving patents and other intellectual property rights in the biotechnology and pharmaceutical industries, as well as administrative proceedings for challenging patents. These include interference, derivation, inter partes review, post-grant review, and reexamination proceedings before the USPTO or oppositions and other comparable proceedings in foreign jurisdictions. Litigation and other legal proceedings relating to intellectual property claims, with or without merit, are unpredictable and generally expensive and time-consuming and, even if resolved in our favor, are likely to divert significant resources from our core business and distract our technical and management personnel from their normal responsibilities. Such litigation or proceedings could substantially increase our operating losses and reduce the resources available for development activities or any future sales, marketing, or distribution activities. We may not have sufficient financial or other resources to adequately conduct such litigation or proceedings. Some of our competitors may be able to sustain the costs of such litigation or proceedings more effectively than we can because of their greater financial resources and more mature and developed intellectual property portfolios. Uncertainties resulting from the initiation and continuation of patent litigation or other proceedings could have a material adverse effect on our ability to enter into or compete in the marketplace. Furthermore, patent reform and changes to patent laws add uncertainty to the possibility of challenge to our patents in the future.
Numerous issued patents and pending patent applications owned by third parties may exist worldwide in the fields in which we are developing our platform technologies and product candidates. We cannot provide any assurances that third-party patent filings that might be enforced against the making, use, or sale of our current product candidates or future products do not exist, which, if they did exist, would result in either an injunction prohibiting our sales, or an obligation to pay royalties on product sales or other forms of compensation to third parties. As the biotechnology and pharmaceutical industries expand and more patents are issued, the risk increases that our product candidates will be subject to claims of infringement of the patent rights of others. Third parties may assert that we infringe their patents or other intellectual property, or that we are otherwise employing their proprietary technology without authorization and may sue us. There may be third-party patent filings of which we are currently unaware with claims, including claims to compositions, formulations, methods of manufacture, or methods of use or treatment, that cover our product candidates. It is also possible that patent filings owned by third parties of which we are aware, but which we do not believe are relevant to our platform technologies, product candidates, or other proprietary technologies we may develop, could be found to be infringed by our product candidate. Because patent applications can take many years to issue, there may be pending patent applications, including those of which we are unaware, that may later result in issued patents that our product candidates may infringe. In addition, third parties, including our competitors in both the United States and abroad, many of which have substantially greater resources and have made substantial investments in patent portfolios and competing technologies, may obtain patents in the future that may prevent, limit, or otherwise interfere with our ability to make, use, and sell our product candidates, and may claim that use of our technologies or the manufacture, use, or sale of our product candidates infringes upon these patents. If any such third-party patents were held by a court of competent jurisdiction to cover our technologies or product candidates, or if we are found to otherwise infringe a third party’s intellectual property rights, the holders of any such patents may be able to block, including by court order, our ability to develop, manufacture, use, sell, or commercialize the applicable product candidate unless we obtain a license under the applicable patents or other intellectual property, or until such patents expire or are finally determined to be held unpatentable, invalid, or unenforceable. Such a license may not be available on commercially reasonable terms or may not be available at all. Even if we were able to obtain a license, it could be non-exclusive, thereby giving our competitors access to the same technologies licensed to us. If we are unable to obtain a necessary license to a third-party patent on commercially reasonable terms, our ability to commercialize our product candidates may be impaired or delayed, which could in turn significantly harm our business.
The pharmaceutical and biotechnology industries have produced a considerable number of patents, and it may not always be clear to industry participants, including us, which patents cover the making, use, or sale of various types of products or methods of use. The scope of patent coverage is subject to interpretation by both administrative bodies and the courts, and the interpretation is not always uniform. If we were sued for patent infringement, we would need to demonstrate that the making, use, or sale of our product candidates, products, or methods either do not infringe the patent claims of the relevant patent or that the patent claims are invalid or unenforceable, and we may not be able to do this. Proving invalidity may be difficult. For example, in the United States, proving invalidity in court requires a showing of clear and convincing evidence to overcome the presumption of validity enjoyed by issued patents, and there is no assurance that a court would invalidate the claims of any such patent. We may not have sufficient resources to bring these actions to a successful conclusion. Even if we are successful in these proceedings, they could cause us to incur substantial costs and divert the time and attention of our management and scientific personnel, which could have a material adverse effect on our business and operations.
Third parties asserting their patent or other intellectual property rights, such as confidential information or trade secrets, against us may seek and obtain injunctive or other equitable relief, which could effectively block our ability to further develop and
108
commercialize our product candidates or force us to cease some of our business operations. Defense against these claims, regardless of their merit, would involve substantial litigation expense and could divert management and other employee resources from our business, cause development delays, and impact our reputation. In the event of a successful claim of infringement against us, we may have to pay substantial damages, including treble damages and attorneys’ fees for willful infringement, obtain one or more licenses from third parties, pay royalties, or redesign our infringing products, which may be impossible to do on a cost-effective basis or require substantial time and monetary expenditure. In that event, we would be unable to further develop and commercialize our product candidates, which could harm our business significantly.
Issued patents and patent applications covering our platform technologies, product candidates, components, or processes in our product development pipeline could be found unpatentable, invalid, or unenforceable if challenged in courts worldwide, including in the United States, or before an administrative body such as the USPTO or comparable foreign authority.
Our issued patents or pending patent applications may be challenged in the courts or patent offices in the United States and abroad. For example, our patent applications may be subject to a third-party pre-issuance submission of prior art to the USPTO, or we may become involved in post-grant review proceedings, opposition or derivation proceedings, reexaminations, or inter partes review proceedings, in the United States or elsewhere, challenging our patent rights or the patent rights of others. An adverse determination in any such proceeding may result in loss of exclusivity or in our patent claims being narrowed, invalidated, held unpatentable, or held unenforceable, in whole or in part, which could limit our ability to exclude others from using or commercializing similar or identical technology and products, or limit the duration of the patent protection of our technology and products, and otherwise no longer protect our product candidates.
In addition, if we or one of our licensing partners initiates legal proceedings against a third party to enforce a patent covering one of platform technologies or one of our product candidates, the defendant could counterclaim that we infringe their patents or that the patent covering our product candidate is invalid or unenforceable, or both. In patent litigation in the United States or abroad, defendant counterclaims alleging invalidity or unenforceability are commonplace, and there are numerous grounds upon which a third party can assert invalidity or unenforceability of a patent, including lack of novelty, obviousness, non-enablement, or insufficient written description or that someone connected with prosecution of the patent withheld relevant information from the USPTO or made a misleading statement during prosecution. Third parties may also raise similar claims before administrative bodies in the United States or abroad, even outside the context of litigation, using post-grant proceedings such as re-examination, inter partes review, post-grant review, opposition, or derivation proceedings. The outcome following legal assertions of unpatentability, invalidity, or unenforceability is unpredictable. In a proceeding before an administrative body, there is a risk that the body will decide that a patent is unpatentable or will be revoked, in whole or in part. In any patent infringement proceeding, there is a risk that a court will decide that a patent of ours is invalid or unenforceable, in whole or in part. In the event of either decision, we would not have the right to stop another party from using the invention at issue. There is also a risk that, even if the validity of such patents is upheld, the court or administrative body will construe the patent’s claims narrowly or decide that we do not have the right to stop the other party from using the invention at issue on the grounds that our patent claims do not cover the invention. The courts could also decide that the other party’s use of our patented technology falls under the safe harbor to patent infringement under 35 U.S.C. §271(e)(1). With respect to the validity and patentability of our patents, for example, we cannot be certain that there is no invalidating prior art of which we, our patent counsel, and the patent offices were unaware during prosecution. If a defendant were to prevail on a legal assertion of invalidity or unenforceability, we would lose at least part, and perhaps all, of the patent protection for the relevant product candidate, which could limit our ability to assert our patents against those parties or other competitors and prevent us from excluding third parties from making, using, or selling similar or competitive products. Any failure to obtain or maintain patent protection with respect to our product candidates could have a material adverse effect on our business, financial condition, results of operations, and prospects.
We may be involved in lawsuits to protect or enforce our patents or other intellectual property or the intellectual property of our licensors, which could be expensive, time-consuming, and unsuccessful.
Competitors may infringe our patents or other intellectual property or the intellectual property of our licensors. To cease such infringement or unauthorized use, we may be required to file patent infringement claims, which can be expensive and time-consuming and could divert the time and attention of our management and scientific personnel. Our pending patent applications cannot be enforced against third parties practicing the technology claimed in such applications unless and until a patent issues from such applications. In addition, in an infringement proceeding or a declaratory judgment action, a court may decide that one or more of our patents is not valid or is unenforceable or may refuse to stop the other party from using the technology at issue on the grounds that our patents do not cover the technology in question. An adverse result in any litigation or defense proceeding could put one or more of our patents at risk of being invalidated, held unenforceable, or interpreted narrowly and could put our patent applications at risk of not issuing. Defense of these claims, regardless of their merit, would involve substantial litigation expense and would require substantial diversion of employee resources from our business.
Interference or derivation proceedings provoked by third parties or brought by the USPTO may be necessary to determine the priority of inventions with respect to, or the correct inventorship of, our patents or patent applications or those of our licensors. An
109
unfavorable outcome could result in a loss of our current patent rights and could require us to cease using the related technology or to attempt to license rights to it from the prevailing party. Our business could be harmed if the prevailing party does not offer us a license on commercially reasonable terms. Litigation, interference, derivation, or other proceedings may result in a decision adverse to our interests and, even if we are successful, may result in substantial costs and distract our management and other employees.
Even if we establish infringement of our or our licensors’ intellectual property, the court may decide not to grant an injunction against further infringing activity and instead award only monetary damages, which may or may not be an adequate remedy. Furthermore, because of the substantial amount of discovery required in connection with intellectual property litigation, there is a risk that some of our confidential information could be compromised by disclosure during this type of litigation. In addition, there could be public announcements of the results of hearings, motions, or other interim proceedings or developments. If securities analysts or investors perceive these results to be negative, the price of our common stock could be substantially adversely affected.
Obtaining and maintaining our patent protection depends on compliance with various procedural, document submission, fee payment, and other requirements imposed by governmental patent agencies, and our patent protection could be reduced or eliminated for non-compliance with these requirements.
The USPTO and foreign governmental patent agencies require compliance with a number of procedural, documentary, fee payment, and other similar requirements during the patent application process. Additionally, periodic maintenance fees on any issued patent must be paid to the USPTO and foreign patent agencies in several stages over the lifetime of the patent. Although an inadvertent lapse, including due to the effect of the COVID-19 pandemic on us or our patent maintenance vendors, can in many cases be cured by payment of a late fee or by other means in accordance with the applicable rules, there are situations in which noncompliance can result in abandonment or lapse of the patent or patent application, resulting in partial or complete loss of patent rights in the relevant jurisdiction. Noncompliance events that could result in a failure to perfect a priority claim, abandonment, or lapse of a patent or patent application include failure to respond to official actions within prescribed time limits, non-payment of fees, and failure to properly legalize and submit formal documents. In any such event, our competitors might be able to enter the market, which would have a material adverse effect on our business.
The terms of our patents may not be sufficient to effectively protect our products and business.
Patents have limited terms, and in many jurisdictions worldwide, including the United States, if all maintenance fees are timely paid, the natural expiration of a patent’s term is generally 20 years after its first effective nonprovisional filing date. Although various extensions may be available, the term of a patent, and the protection it affords, is limited. Given the significant amount of time required for the development, testing, and regulatory review of new product candidates, patents protecting such product candidates might expire before or shortly after such product candidates are commercialized. Further, if we encounter delays in our clinical trials, the period of time during which we could market our product candidates under patent protection would be reduced. Even if patents covering our product candidates are obtained, once the patent life has expired for a product, we may be open to competition from biosimilar or generic therapies. As a result, our patent portfolio may not provide us with sufficient rights to exclude others from commercializing product candidates similar or identical to ours. Our patents issued as of February, 2022, have terms expected to expire on dates ranging from 2023 to 2040, subject to any patent extensions that may be available for such patents. If patents are issued on our patent applications pending as of February 2022, the resulting patents are projected to expire on dates ranging from 2023 to 2043. In addition, although upon issuance in the United States a patent’s term can be increased based on certain delays caused by the USPTO, this increase can be reduced or eliminated based on certain delays caused by the patent applicant during patent prosecution. A patent term extension based on regulatory delay may also be available in the United States and in certain other foreign jurisdictions. However, in the United States, only a single patent can be extended for each marketing approval, and any patent can be extended only once, for a single product. Moreover, the scope of protection during the period of the patent term extension in the United States does not extend to the full scope of the patent’s claim, but instead only as to the scope of the product as approved. The laws governing analogous patent term extensions in foreign jurisdictions vary widely and many differ from the process in the United States. Additionally, we may not receive an extension of patent term if we fail to exercise due diligence during the testing phase or regulatory review process, fail to apply within applicable deadlines, fail to apply prior to expiration of relevant patents, or otherwise fail to satisfy applicable requirements. If we are unable to obtain patent term extension, or the term of any such extension is less than we request, the period during which we will have the right to exclude others from using the patent rights will be shortened. Our competitors may be able to obtain approval of competing products following our patent expiration and take advantage of our investment in development and clinical trials by referencing our clinical and preclinical data to launch a biosimilar product earlier than might otherwise be the case, which could reduce our revenue, possibly materially. In general, if we do not have sufficient patent term to protect our platform technologies and product candidates, our business and results of operations will be adversely affected.
We may be subject to claims challenging the inventorship or ownership of our patent and other intellectual property rights.
We may be subject to claims that former employees, collaborators, or other third parties have an ownership interest in our patents or other intellectual property, including as a result of being an inventor or co-inventor. In the United States, the failure to name
110
the proper inventors on a granted patent can result in the patent being unenforceable. Inventorship disputes may arise from conflicting views regarding the contributions made to an invention by the individuals named as inventors, the effects of foreign laws where foreign nationals are involved in the development of the subject matter of the patent, conflicting obligations of third parties involved in developing our product candidates, or as a result of questions regarding co-ownership of potential joint inventions. For example, inventorship disputes may arise from conflicting obligations of consultants or others who are involved in developing our platform technologies or product candidates or related intellectual property. Alternatively, or additionally, we may enter into agreements to clarify the scope of our rights in such intellectual property. Litigation may be necessary to defend against claims challenging or relating to inventorship and ownership of intellectual property rights. If we fail in defending any such claims, in addition to paying monetary damages, we may lose valuable intellectual property rights, such as exclusive ownership of, or the right to use, valuable intellectual property. Such an outcome could have a material adverse effect on our business. Even if we are successful in defending against such claims, litigation could result in substantial costs and distract management and other employees.
We or our licensors may have relied on third-party consultants or collaborators or on funds from third parties, such as the United States government, such that we or our licensors are not the sole and exclusive owners of the patents that we own or that we have in-licensed. If other third parties have ownership rights or other rights to our patents, including in-licensed patents, they may be able to license such patents to our competitors, and our competitors could make, use, or sell competing products and technology. This could have a material adverse effect on our competitive position, business, financial conditions, results of operations, and prospects.
In addition, while it is our policy to require our employees and contractors who may be involved in the conception or development of intellectual property to execute agreements assigning such intellectual property to us, we may be unsuccessful in executing such an agreement with each party who, in fact, conceives or develops intellectual property that we regard as our own. The assignment of intellectual property rights may not be self-executing, or the assignment agreements may be breached, and we may be forced to bring claims against third parties, or defend claims that they may bring against us, to determine the ownership of what we regard as our intellectual property. As described elsewhere in these Risk Factors, such claims could be expensive and time-consuming to litigate or defend and could divert the time and attention of our management and scientific personnel, which could have a material adverse effect on our business, financial condition, results of operations, and prospects.
We may be subject to claims that our employees, consultants, or independent contractors have wrongfully used or disclosed confidential information of third parties.
We have received confidential and proprietary information from third parties. In addition, we employ individuals who were previously employed by other organizations, including at other biotechnology or pharmaceutical companies or at academic institutions. We may be subject to claims that we or our employees, consultants, or independent contractors have inadvertently or otherwise improperly used or disclosed confidential information of these third parties or our employees’ former employers or that we caused an employee to breach the terms of his or her non-competition or non-solicitation agreement. Litigation may be necessary to defend against these claims. Even if we are successful in defending against these claims, litigation could cause us to incur substantial costs and distract our management and employees. If our defenses to these claims fail, in addition to requiring us to pay monetary damages, a court could prohibit us from using technologies or features that are essential to our product candidates, if such technologies or features are found to incorporate or be derived from the trade secrets or other proprietary information of these third parties or former employers. Moreover, any such litigation or the threat thereof may adversely affect our reputation and our ability to form strategic alliances or sublicense our rights to collaborators, engage with scientific advisors, or hire employees or consultants, each of which would have an adverse effect on our business, results of operations, and financial condition.
Our internal computer systems, or those used by our third-party research institution collaborators, CROs, CDMOs, or other contractors or consultants, may fail or suffer security breaches.
We are increasingly dependent upon information technology systems, infrastructure, and data to operate our business. In the ordinary course of business, we collect, store, and transmit confidential information (including intellectual property, proprietary business information, and personal information). It is critical that we do so in a secure manner to maintain the confidentiality and integrity of such confidential information. We also have outsourced elements of our operations to third parties, and as a result we manage a number of third-party vendors who have access to our confidential information, including third-party vendors of information technology and data security systems and services. While we generally have agreements requiring such vendors to use industry standard practices for data security, we have no operational control over them.
Despite the implementation of security measures (including edge technology designed to identify and protect our network from infiltration by third-party systems), our internal computer systems and those of our CROs, CDMOs, and other contractors and consultants as well as third-party vendors of information technology and data security systems and services are vulnerable to damage and interruptions from security breaches, computer viruses, ransomware, fraud, and similar incidents involving the loss or unauthorized access of confidential information. One third-party vendor that experienced such an incident is SolarWinds Corporation (SolarWinds), a provider of information technology monitoring and management products and services, including its Orion Platform
111
products, which are used by over 30,000 businesses, including ours. SolarWinds experienced a cyberattack that appears likely to be the result of a supply chain attack by an outside nation state. SolarWinds has stated that, as a result of the attack, software updates related to its Orion Platform products delivered between March and June 2020 included vulnerabilities, and that its investigation is ongoing. Since being notified of the attack, we have taken steps to mitigate the vulnerabilities identified within the Orion Platform products. We also conducted investigations to determine the extent to which our confidential information was accessed, lost, or stolen as a result of this cyberattack on SolarWinds and concluded that our confidential information was not materially accessed, lost, or stolen as a result of the cyberattack. We continue to monitor our systems and upgrade our security capabilities in order to mitigate risk. However, any access, loss, or theft of our confidential information in connection with a future cyberattack could have a material adverse effect on our business.
Threats involving the misuse of access our network, systems, and information by our current or former employees, contractors, vendors, or partners, whether intentional or unintentional, also pose a risk to the security of our network, systems, and information and data. For example, we are subject to the risk that employees may inadvertently share confidential information with unintended third parties, or that departing employees may take, or create their own information based on, our confidential information upon leaving the company. In addition, any such insiders may be the victims of social engineering attacks that enable third parties to access our network, systems, and information using an authorized person’s credentials. We and our network, systems, and information are also vulnerable to malicious acts by insiders, including leaking, modifying, or deleting confidential information, or performing other acts that could materially interfere with our operations and business. While we provide regular training to our employees regarding cybersecurity threats and best practices, we cannot ensure that such training or other efforts will prevent unauthorized access to or sabotage of our network, systems, and information.
While we have not, to our knowledge, experienced any material system failure, accident, or security breach to date, because techniques used to obtain unauthorized access to or to sabotage systems are constantly evolving and generally are not recognized until they are launched against a target, we cannot be sure that our continued data protection efforts and investment in information technology will prevent future significant breakdowns, data leakages, breaches in our systems or the systems of our third party contractors and collaborators, or other cyber incidents that could have a material adverse effect upon our reputation, business, operations, or financial condition. If such an event were to occur, it could materially disrupt our operations and programs, and the development of our product candidates could be delayed. For example, the loss of or inability to access clinical trial data for our product candidates could result in delays in further development of our product candidates and in our regulatory, marketing approval, and commercialization efforts and significantly increase our costs to recover or reproduce the data. Furthermore, significant disruptions or security breaches of our internal information technology systems or our third party contractors and collaborators’ information technology systems could result in the loss, misappropriation, or unauthorized access, use, or disclosure of, or the prevention of access to, our confidential information (including trade secrets or other intellectual property, proprietary business information, and personal information), which could also result in financial, legal, business, and reputational harm to us. Any such event that leads to unauthorized access, use, or disclosure of personal information, including personal information regarding our clinical trial subjects or employees, could delay further development and commercialization of our product candidates, harm our reputation directly, require us to comply with federal or state breach notification laws and foreign law equivalents, subject us to mandatory corrective action, and otherwise subject us to liability under laws and regulations that protect the privacy and security of personal information. As a result, we could incur significant legal and financial exposure and reputational damages that could have a material adverse effect on our business.
In addition, we have and will continue to enter into collaboration, license, contract research and manufacturing relationships with organizations that operate in certain countries that are at heightened risk of theft of technology, data, and intellectual property through direct intrusion by private parties or foreign actors, including those affiliated with or controlled by state actors. If any theft affects our technology, data, or intellectual property, our efforts to protect and enforce our intellectual property rights around the world may be inadequate to obtain a significant commercial advantage from our intellectual property, and we may be at heightened risk of losing our proprietary intellectual property rights around the world, including outside of such countries, to the extent such theft or intrusion destroys the proprietary nature of our intellectual property.
If our trademarks and trade names are not adequately protected, then we may not be able to build name recognition in our markets of interest and our business may be adversely affected.
Our current or future trademarks or trade names may be challenged, infringed, circumvented, or declared generic or descriptive, or may be determined to infringe on other marks. We may not be able to protect our rights to these trademarks and trade names or may be forced to stop using these trademarks or trade names, which we need for name recognition by potential partners or customers in our markets of interest. During trademark registration proceedings, we may receive rejections of our applications by the USPTO or in other foreign jurisdictions. Although we would be given an opportunity to respond to those rejections, we may be unable to overcome such rejections. In addition, in the USPTO and in comparable agencies in many foreign jurisdictions, third parties are given an opportunity to oppose pending trademark applications and to seek to cancel registered trademarks. Opposition or cancellation proceedings may be filed against our trademarks, and our trademarks may not survive such proceedings. If we are unable to establish
112
name recognition based on our trademarks and trade names, we may not be able to compete effectively, and our business may be adversely affected. We may license our trademarks and trade names to collaborators or to third parties, such as distributors. Though these license agreements may provide guidelines for how our trademarks and trade names may be used, a breach of these agreements or misuse of our trademarks and tradenames by our licensees may jeopardize our rights in or diminish the goodwill associated with our trademarks and trade names.
Moreover, any name we may propose to use as a trade name for any of our product candidates in the United States must be approved by the FDA, regardless of whether we have applied to register it, as a trademark. Similar requirements exist in Europe. The FDA typically conducts a review of proposed product names, including an evaluation of potential for confusion with other product names. If the FDA or a comparable foreign regulatory authority objects to any of our proposed proprietary product names, we may be required to expend significant additional resources in an effort to identify a suitable substitute name that would be registerable under applicable trademark laws, not infringe the existing rights of third parties and be acceptable to the FDA. Furthermore, in many countries, owning and maintaining a trademark registration may not provide an adequate defense against a subsequent infringement claim asserted by the owner of a senior trademark. At times, competitors or other third parties may adopt trademarks or trade names similar to ours, thereby impeding our ability to build brand identity and possibly leading to market confusion. In addition, there could be potential trademark or trade name infringement claims brought by owners of other registered trademarks or trademarks that incorporate variations of our registered or unregistered trademarks or trade names. If we assert trademark infringement claims, a court may determine that the marks we have asserted are invalid or unenforceable, or that the party against whom we have asserted trademark infringement has superior rights to the marks in question. In this case, we could ultimately be forced to cease use of such trademarks.
Changes in United States patent law could diminish the value of patents in general, thereby impairing our ability to protect our products.
As is the case with other biopharmaceutical companies, our success is heavily dependent on intellectual property, particularly patent rights. Obtaining and enforcing patents in the biopharmaceutical industry involves both technological and legal complexity and is therefore costly, time-consuming, and inherently uncertain. Changes in either the patent laws or interpretation of the patent laws in the United States could increase the uncertainties and costs associated with protection of, and may diminish our ability to protect, our inventions and obtain, maintain, and enforce our intellectual property rights and, more generally, could adversely affect the value of our intellectual property or narrow the scope of our owned and licensed patents. Recent patent reform legislation in the United States and other countries, including the Leahy-Smith Act signed into law on September 16, 2011, could increase uncertainties and costs surrounding the prosecution of our patent applications and the enforcement or defense of our issued patents. The Leahy-Smith Act introduced a number of significant changes to United States patent law. These include provisions that affect the way patent applications are prosecuted, redefine prior art, and provide more efficient and cost-effective avenues for competitors to challenge patents. These include allowing third-party submission of prior art to the USPTO during patent prosecution and additional procedures to attack patents by USPTO-administered post-grant proceedings, including post-grant review, inter partes review, and derivation proceedings. In addition, under the Leahy-Smith Act, the United States transitioned to a first inventor to file system for filings made after March 2013 in which, assuming that the other statutory requirements are met, the first inventor to file a patent application will be entitled to the patent on an invention regardless of whether a third-party was the first to invent the claimed invention. A third party that files a patent application in the USPTO after March 2013, but before we file an application covering the same invention, could therefore be awarded a patent covering an invention of ours even if we had made the invention before it was made by such third party. This requires us to be cognizant of the time from invention to filing of a patent application. Since patent applications in the United States and most other countries are confidential for a period of time after filing or until issuance, we cannot be certain that we or our licensors were the first to either (i) file any patent application related to our platform technologies, product candidates, and other proprietary technologies we may develop or (ii) invent any of the inventions claimed in our or our licensor’s patents or patent applications. Even where we have a valid and enforceable patent, we may not be able to exclude others from practicing the claimed invention where the other party can show that they used the invention in commerce before our filing date or the other party benefits from a compulsory license. The Leahy-Smith Act and its implementation could increase the uncertainties and costs surrounding the prosecution of our patent applications and the enforcement or defense of our issued patents, all of which could have a material adverse effect on our business, financial condition, results of operations, and prospects.
In addition, recent United States Supreme Court rulings have narrowed the scope of patent protection available in certain circumstances and weakened the rights of patent owners in certain situations. In addition to increasing uncertainty with regard to our ability to obtain patents in the future, this combination of events has created uncertainty with respect to the value of patents once obtained. Depending on decisions by Congress, the federal courts, the USPTO, and the relevant law-making bodies in other countries, the laws and regulations governing patents could change in unpredictable ways that would weaken our ability to obtain new patents or to enforce our existing patents and patents that we might obtain in the future. For example, in the 2013 case Assoc. for Molecular Pathology v. Myriad Genetics, Inc., the United States Supreme Court held that certain claims to naturally occurring substances are not
113
patentable. Although we do not believe that any of the patents owned or licensed by us will be found invalid based on this decision, we cannot predict how future decisions by Congress, the federal courts or the USPTO may impact the value of our patents.
Risks Related to Our Regulatory Environment
The development and commercialization of biopharmaceutical products is subject to extensive regulation, and the regulatory approval processes of the FDA and comparable foreign authorities are lengthy, time-consuming, and inherently unpredictable. If we are unable to obtain regulatory approval for our product candidates on a timely basis, or at all, our business will be substantially harmed.
The clinical development, manufacturing, labeling, packaging, storage, recordkeeping, advertising, promotion, export, import, marketing, distribution, adverse event reporting, including the submission of safety and other post-marketing information and reports, and other activities we may engage in relating to our product candidates are subject to extensive regulation. In the United States, marketing approval of biologics requires the submission of a BLA to the FDA, and we will not be permitted to market any product candidate in the United States until the FDA has approved the BLA for that product candidate. A BLA must be supported by extensive clinical and preclinical data, as well as extensive information regarding pharmacology, chemistry, manufacturing, and controls. Outside the United States, many comparable foreign regulatory authorities employ similar approval processes.
To date, we have not submitted a BLA to the FDA or similar regulatory approval filings to comparable foreign authorities for any product candidate, and we cannot be certain that any of our product candidates will receive regulatory approval once a BLA or similar application has been submitted. Obtaining approval of a BLA can be a lengthy, expensive, and uncertain process, and as a company we have no experience with the preparation and submission of a BLA or any other application for marketing approval. Further, the FDA has not yet granted approval for a therapeutic derived from stem cells, which we believe may increase the complexity, uncertainty, and length of the regulatory approval process for certain of our product candidates derived from our ex vivo cell engineering platform. In addition, the FDA has the authority to require a REMS plan as part of a BLA approval or after BLA approval, which may impose further requirements or restrictions on the distribution or use of an approved biologic, such as limiting prescribing to certain physicians or medical centers that have undergone specialized training, limiting treatment to patients who meet certain safe-use criteria, and requiring treated patients to enroll in a registry.
Our product candidates could fail to receive regulatory approval for many reasons, including the following:
|
|
• |
the FDA or comparable foreign regulatory authorities may disagree with the design or implementation of our clinical trials; |
|
|
• |
we may be unable to demonstrate to the satisfaction of the FDA or comparable foreign regulatory authorities that a product candidate is safe and effective for its proposed indication; |
|
|
• |
the results of clinical trials may not meet the level of statistical significance required by the FDA or comparable foreign regulatory authorities for approval; |
|
|
• |
we may be unable to demonstrate that a product candidate’s clinical and other benefits outweigh its safety risks; |
|
|
• |
the FDA or comparable foreign regulatory authorities may disagree with our interpretation of data from preclinical studies or clinical trials; |
|
|
• |
the data collected from clinical trials may not be sufficient to support the submission of a BLA or other comparable foreign submission or to obtain regulatory approval in the United States or elsewhere, or regulatory authorities may not accept a submission due to, among other reasons, the content or formatting of the submission; |
|
|
• |
the FDA or comparable foreign regulatory authorities may fail to approve our manufacturing processes or facilities or those of third-party manufacturers with which we contract for clinical and commercial product supply; and |
|
|
• |
the approval policies or regulations of the FDA or comparable foreign regulatory authorities may change in a manner that renders our clinical data insufficient for approval, including, for example, as a result of positive or negative data from third parties regarding other products or product candidates. |
The lengthy approval process, as well as the unpredictability of future clinical trial results, may prevent us from obtaining regulatory approval to market any of our product candidates, which would significantly harm our business, results of operations, and prospects. The FDA and comparable foreign regulatory authorities have substantial discretion in the approval process and in determining whether and when regulatory approval will be granted for any of our product candidates. For example, regulatory authorities in various jurisdictions have in the past had, and may in the future have, differing requirements for, interpretations of, and opinions on our preclinical and clinical data. As a result, we may be required to conduct additional preclinical studies, alter our proposed clinical trial designs, or conduct additional clinical trials to satisfy the regulatory authorities in each of the jurisdictions in which we hope to conduct clinical trials and develop and market our products, if approved. Further, even if we believe the data
114
collected from clinical trials of our product candidates are promising, such data may not be sufficient to support approval by the FDA or any comparable foreign regulatory authority.
In addition, even if we were to obtain approval for any of our product candidates, regulatory authorities may grant such approval for fewer or more limited indications than we request, may not approve the price we intend to charge for such product, may grant approval contingent on the performance of costly post-marketing clinical trials, or may approve such product with a label that does not include the labeling claims necessary or desirable for the successful commercialization of that product. Notably, to date, the FDA has required that any patient receiving a gene therapy be followed for 15 years post-treatment. This post-treatment follow-up increases the cost and complexity of commercializing gene therapy products. Any of the foregoing scenarios could materially harm the commercial prospects for our product candidates.
Even if our product candidates obtain regulatory approval, we will be subject to ongoing obligations and continued regulatory review, which may result in significant additional expense. Additionally, our product candidates, if approved, could be subject to labeling and other restrictions and market withdrawal, and we may be subject to penalties if we fail to comply with regulatory requirements or experience unanticipated problems with our products.
If the FDA or a comparable foreign regulatory authority approves any of our product candidates, the manufacturing processes, testing, labeling, packaging, distribution, import, export, adverse event reporting, storage, advertising, promotion, and recordkeeping for the product will be subject to extensive and ongoing regulatory requirements. These requirements include submission of safety and other post-marketing information and reports, registration, as well as continued compliance with cGMP and GCP regulations for any clinical trials that we conduct post-approval, all of which may result in significant expense and limit our ability to commercialize such products. In addition, any regulatory approvals that we receive for our product candidates may also be subject to limitations on the approved indicated uses for which the product may be marketed or to the conditions of approval, or contain requirements for potentially costly post-marketing testing, including Phase 4 clinical trials, and surveillance to monitor the safety and efficacy of the approved product.
Manufacturers and manufacturers’ facilities are required to comply with extensive FDA and comparable foreign regulatory authority requirements, including ensuring that quality control and manufacturing procedures conform to cGMP regulations, as well as, for the manufacture of certain of our product candidates, the FDA’s cGTPs for the use of human cellular and tissue products to prevent the introduction, transmission, or spread of communicable diseases. As such, we and our contract manufacturers will be subject to continual review and inspections to assess compliance with cGMP and cGTP regulations and adherence to commitments made in any approved marketing application. Accordingly, we and third parties that we engage or with which we conduct business must continue to expend time, money, and effort in all areas of regulatory compliance, including manufacturing, quality control, and distribution.
If there are changes in the application of legislation or regulatory policies, or if problems are discovered with an approved product or our manufacture of such a product, or if we or one of our distributors, licensees, or co-marketers fails to comply with regulatory requirements, United States and foreign regulatory authorities could take various actions. These may include issuing warning letters or untitled letters, imposing fines on us, imposing restrictions on the applicable product or its manufacture, or requiring us to recall or remove the product from the market. Regulatory authorities could also suspend or withdraw our marketing authorizations, which could require us to conduct additional clinical trials, change our product labeling, or submit additional applications for marketing authorization. If any of these events were to occur, our ability to sell such product may be impaired, and we may incur substantial additional expense to comply with regulatory requirements, which could materially adversely affect our business, financial condition, and results of operations.
In addition, if we obtain approval for any of our product candidates, our product labeling, advertising, and promotion will be subject to regulatory requirements and continuing regulatory review. In the United States, the FDA and the Federal Trade Commission (FTC) strictly regulate the promotional claims that may be made about pharmaceutical products to ensure that any claims about such products are consistent with regulatory approvals, not misleading or false in any particular way, and adequately substantiated by clinical data. The promotion of a drug product in a manner that is false, misleading, unsubstantiated, or for unapproved (or off-label) uses may result in enforcement letters, inquiries and investigations, and civil and criminal sanctions by the FDA or the FTC. In particular, a product may not be promoted for uses that are not approved by the FDA, as reflected in the product’s approved labeling. If we receive marketing approval for a product candidate, physicians may nevertheless prescribe it to their patients in a manner that is inconsistent with the approved label. If we are found to have promoted such off-label uses, we may be subject to significant liability. The FDA and other agencies actively enforce the laws and regulations prohibiting the promotion of off-label uses, and a company that is found to have improperly promoted off-label uses may be subject to significant sanctions and false claims litigation under federal and state statutes, which can lead to consent decrees, civil monetary penalties, restitution, criminal fines and imprisonment, and exclusion from participation in Medicare, Medicaid, and other federal and state healthcare programs. The federal government has levied large civil and criminal fines against companies for alleged improper promotion and has enjoined several companies from
115
engaging in off-label promotion. The FDA has also requested that companies enter into consent decrees or permanent injunctions under which specified promotional conduct is changed or curtailed.
If a regulatory agency discovers previously unknown problems with a product, such as adverse events of unanticipated severity or frequency, or problems with the facility where the product is manufactured, or disagrees with the promotion, marketing, or labeling of a product, such regulatory agency may impose restrictions on that product or us, including requiring withdrawal of the product from the market. If we fail to comply with applicable regulatory requirements, a regulatory agency or enforcement authority may, among other things:
|
|
• |
issue warning letters; |
|
|
• |
issue, or require us to issue, safety-related communications, such as safety alerts, field alerts, “Dear Doctor” letters to healthcare professionals, or import alerts; |
|
|
• |
impose civil or criminal penalties; |
|
|
• |
suspend, limit, or withdraw regulatory approval; |
|
|
• |
suspend any of our preclinical studies and clinical trials; |
|
|
• |
refuse to approve pending applications or supplements to approved applications submitted by us; |
|
|
• |
impose restrictions on our operations, including requiring us to close our and our CDMOs’ facilities; or |
|
|
• |
seize or detain products, refuse to permit the import or export of products, or require us to conduct a product recall. |
Any government investigation of alleged violations of law could require us to expend significant time and resources in response and could generate negative publicity. Any failure to comply with ongoing regulatory requirements may significantly and adversely affect our ability to commercialize and generate revenue from our products, if approved. If regulatory sanctions are applied or if regulatory approval is withdrawn, the value of our company and our operating results will be adversely affected.
Moreover, the policies of the FDA and of comparable foreign regulatory authorities may change and additional government regulations may be enacted that could prevent, limit, or delay regulatory approval of our product candidates. We cannot predict the likelihood, nature, or extent of government regulation that may arise from future legislation or administrative or executive action, either in the United States or abroad. If we are slow or unable to adapt to changes in existing requirements or the adoption of new requirements or policies, or if we are not able to maintain regulatory compliance, we may be subject to enforcement action and we may not achieve or sustain profitability.
Disruptions at the FDA and other government agencies caused by funding shortages or global health concerns could hinder their ability to hire, retain, or deploy key leadership and other personnel, or otherwise prevent new or modified products from being developed, approved, or commercialized in a timely manner or at all, which could negatively impact our business.
The ability of the FDA to review and approve new products may be affected by a variety of factors, including government budget and funding levels, statutory, regulatory, and policy changes, the FDA’s ability to hire and retain key personnel, including personnel with the expertise necessary to evaluate product candidates such as ours, and accept the payment of user fees, and other events that may otherwise affect the FDA’s ability to perform routine functions. Average review times at the FDA have fluctuated in recent years. Moreover, these and other factors have increased the uncertainties associated with interpreting the FDA’s guidance and predicting its areas of focus and responses to various issues. In addition, government funding of other government agencies that fund research and development activities is subject to the political process, which is inherently fluid and unpredictable. Disruptions at the FDA and other agencies may also extend the time necessary for new biologics or modifications to licensed biologics to be reviewed or approved by necessary government agencies, which would adversely affect our business. For example, over the last several years, the United States government has shut down several times and certain regulatory agencies, such as the FDA, have had to furlough critical FDA employees and stop critical activities.
Separately, in response to the ongoing COVID-19 pandemic, in March 2020, the FDA announced its intention to postpone most inspections of foreign manufacturing facilities, and on March 18, 2020, the FDA temporarily postponed routine surveillance inspections of domestic manufacturing facilities. Subsequently, in July 2020, the FDA resumed certain on-site inspections of domestic manufacturing facilities subject to a risk-based prioritization system. The FDA utilized this risk-based assessment system to assist in determining when and where it was safest to conduct prioritized domestic inspections. Additionally, on April 15, 2021, the FDA issued a guidance document in which the FDA described its plans to conduct voluntary remote interactive evaluations of certain drug manufacturing facilities and clinical research sites, among other facilities. According to the guidance, the FDA may request such remote interactive evaluations where the FDA determines that remote evaluation would be appropriate based on mission needs and travel limitations. In May 2021, the FDA outlined a detailed plan to move toward a more consistent state of inspectional operations,
116
and in July 2021, the FDA resumed standard inspectional operations of domestic facilities and was continuing to maintain this level of operation as of September 2021. More recently, the FDA has continued to monitor and implement changes to its inspectional activities to ensure the safety of its employees and those of the firms it regulates as it adapts to the evolving COVID-19 pandemic. Regulatory authorities outside the United States may adopt similar restrictions or other policy measures in response to the ongoing COVID-19 pandemic. If a prolonged government shutdown occurs, or if global health concerns continue to prevent the FDA or comparable foreign regulatory authorities from conducting their regular inspections, reviews, or other regulatory activities, it could significantly impact the ability of the FDA or comparable foreign regulatory authorities to timely review and process our regulatory submissions, which could have a material adverse effect on our business.
Our business operations and current and future relationships with healthcare professionals, principal investigators, consultants, vendors, customers, and third-party payors in the United States and elsewhere are subject to applicable anti-kickback, fraud and abuse, false claims, physician payment transparency, and other healthcare laws and regulations, which could expose us to substantial penalties, contractual damages, reputation harm, administrative burdens, and diminished profits.
Healthcare providers, healthcare facilities and institutions, physicians, and third-party payors in the United States and elsewhere will play a primary role in the recommendation and prescription of any product candidates for which we may obtain marketing approval. Our current and future arrangements with healthcare professionals, healthcare facilities and institutions, principal investigators, consultants, customers, and third-party payors may expose us to broadly applicable fraud and abuse and other healthcare laws, including the federal Anti-Kickback Statute and the federal False Claims Act, that may constrain the business or financial arrangements and relationships through which we research, sell, market, and distribute any product candidates for which we obtain marketing approval. In addition, we may be subject to physician payment transparency laws and regulation by the federal and state governments and by foreign jurisdictions in which we conduct our business. The applicable federal, state, and foreign healthcare laws that affect our ability to operate include, but are not limited to, the following:
|
|
• |
the federal Anti-Kickback Statute, which prohibits, among other things, persons or entities from knowingly and willfully soliciting, offering, receiving, or providing any remuneration (including any kickback, bribe, or certain rebate), directly or indirectly, overtly or covertly, in cash or in kind, to induce or reward, or in return for, either the referral of an individual for, or the purchase, lease, order, or recommendation of, any good, facility, item, or service, for which payment may be made, in whole or in part, under any United States federal healthcare program, such as Medicare and Medicaid. The term “remuneration” has been broadly interpreted to include anything of value, including stock options. The federal Anti-Kickback Statute has also been interpreted to apply to arrangements between pharmaceutical manufacturers on the one hand and prescribers, purchasers, and formulary managers on the other hand. There are a number of statutory exceptions and regulatory safe harbors protecting some common activities from prosecution, but the exceptions and safe harbors are drawn narrowly and require strict compliance in order to offer protection. Any arrangements with prescribers must be for bona fide services and compensated at fair market value. A person or entity does not need to have actual knowledge of the statute or specific intent to violate it in order to have committed a violation; |
|
|
• |
the United States federal civil and criminal false claims laws, including the civil False Claims Act, which can be enforced by private citizens on behalf of the United States federal government through civil whistleblower or qui tam actions, and the federal civil monetary penalties law which prohibit, among other things, individuals or entities from knowingly presenting, or causing to be presented, to the United States federal government, claims for payment or approval that are false or fraudulent, knowingly making, using, or causing to be made or used, a false record or statement material to a false or fraudulent claim, or from knowingly making a false statement to avoid, decrease, or conceal an obligation to pay money to the United States federal government. Pharmaceutical manufacturers can cause false claims to be presented to the United States federal government by, among other things, engaging in impermissible marketing practices, such as the off-label promotion of a product for an indication for which it has not received FDA approval. Further, pharmaceutical manufacturers can be held liable under the civil False Claims Act even when they do not submit claims directly to government payors if they are deemed to “cause” the submission of false or fraudulent claims. In addition, the government may assert that a claim including items and services resulting from a violation of the federal Anti-Kickback Statute constitutes a false or fraudulent claim for purposes of the civil False Claims Act; |
|
|
• |
the federal Health Insurance Portability and Accountability Act of 1996 (HIPAA), which imposes criminal and civil liability for, among other things, knowingly and willfully executing, or attempting to execute, a scheme to defraud any healthcare benefit program, or knowingly and willfully falsifying, concealing, or covering up a material fact or making any materially false statement, in connection with the delivery of, or payment for, healthcare benefits, items, or services. Similar to the federal Anti-Kickback Statute, a person or entity does not need to have actual knowledge of the healthcare fraud statute implemented under HIPAA or specific intent to violate it in order to have committed a violation; |
|
|
• |
the United States federal Food, Drug, and Cosmetic Act (the FDCA), which prohibits, among other things, the adulteration or misbranding of drugs, biologics, and medical devices; |
117
|
|
• |
the United States Public Health Service Act, which prohibits, among other things, the introduction into interstate commerce of a biological product unless a biologics license is in effect for that product; |
|
|
• |
the United States Physician Payments Sunshine Act and its implementing regulations, which requires, among other things, certain manufacturers of drugs, devices, biologics, and medical supplies that are reimbursable under Medicare, Medicaid, or the Children’s Health Insurance Program, with specific exceptions, to report annually to the Centers for Medicare and Medicaid Services (CMS) information related to certain payments and other transfers of value to physicians, as defined by statute, certain non-physician practitioners ( including physician assistants, nurse practitioners, clinical nurse specialists, certified nurse anesthetists, anesthesiologist assistants, and certified nurse-midwives), and teaching hospitals, as well as ownership and investment interests held by such physicians and their immediate family members; |
|
|
• |
analogous United States state laws and regulations, including: state anti-kickback and false claims laws, which may apply to our business practices, including research, distribution, sales and marketing arrangements, and claims involving healthcare items or services reimbursed by any third-party payor, including private insurers; state laws that require pharmaceutical companies to comply with the pharmaceutical industry’s voluntary compliance guidelines and the relevant compliance guidance promulgated by the United States federal government, or otherwise restrict payments that may be made to healthcare providers and other potential referral sources; state laws and regulations that require drug manufacturers to file reports relating to pricing and marketing information, which requires tracking gifts and other remuneration and items of value provided to healthcare professionals and entities; and state and local laws requiring the registration of pharmaceutical sales representatives; and |
|
|
• |
similar healthcare laws and regulations in foreign jurisdictions, including reporting requirements detailing interactions with and payments to healthcare providers. |
Ensuring that our internal operations and future business arrangements with third parties comply with applicable healthcare laws and regulations will involve substantial costs. Efforts to ensure that our business arrangements will comply with applicable healthcare laws may involve substantial costs. We have entered into consulting and scientific advisory board arrangements with physicians and other healthcare providers, including some who could influence the use of our product candidates, if approved. Compensation under some of these arrangements includes the provision of stock or stock options in addition to cash consideration. Because of the complex and far-reaching nature of these laws, it is possible that governmental authorities could conclude that our payments to physicians may not be fair market value for bona fide services or that our business practices do not comply with current or future statutes, regulations, agency guidance or case law involving applicable fraud and abuse or other healthcare laws and regulations. If our operations are found to be in violation of any of the laws described above or any other governmental laws and regulations that may apply to us, we may be subject to significant penalties, including civil, criminal, and administrative penalties, damages, fines, exclusion from government-funded healthcare programs, such as Medicare and Medicaid, or similar programs in other countries or jurisdictions, integrity oversight and reporting obligations to resolve allegations of noncompliance, disgorgement, imprisonment, contractual damages, reputational harm, diminished profits, and the curtailment or restructuring of our operations. If any of the physicians or other providers or entities with whom we expect to do business are found to not be in compliance with applicable laws, they may be subject to criminal, civil or administrative sanctions, including exclusions from government funded healthcare programs and imprisonment, which could affect our ability to operate our business. Further, defending against any such actions can be costly, time-consuming and may require significant personnel resources. Therefore, even if we are successful in defending against any such actions that may be brought against us, our business may be impaired.
Our employees, independent contractors, principal investigators, consultants, commercial partners, and vendors may engage in misconduct or other improper activities, including noncompliance with regulatory standards and requirements.
We are exposed to the risk of fraud and other misconduct committed by our personnel and third parties that we engage or with which we collaborate in the course of our operations. It is not always possible to identify and deter misconduct or business noncompliance by our employees, consultants, and other agents, and we cannot ensure that precautions we take to detect and prevent inappropriate conduct, including our compliance controls, policies, and procedures, will in every instance protect us or be effective in controlling unknown or unmanaged risks or losses or in protecting us from governmental investigations or other actions or lawsuits stemming from acts committed by our employees, agents, contractors, or collaborators that would violate the laws or regulations of the jurisdictions in which we operate, including employment, foreign corrupt practices, trade restrictions and sanctions, environmental, competition, and patient privacy and other data privacy and protection laws and regulations. Misconduct by employees, independent contractors, principal investigators, consultants, commercial partners, and vendors could include failures to comply with FDA regulations, provide accurate information to the FDA, comply with manufacturing standards we may establish, report financial information or data accurately, comply with federal and state healthcare fraud and abuse laws and regulations, including prohibitions on pricing, discounting, labeling, marketing and promotion, sales commission, customer incentive programs, and other business arrangements, or disclose unauthorized activities to us. Misconduct by our employees, independent contractors, principal investigators, consultants, commercial partners, and vendors could also involve the improper use of information obtained in the course of clinical trials, which could result in regulatory sanctions and serious harm to our reputation. It is not always possible to identify and
118
deter misconduct, and the precautions we take to detect and prevent this activity may not be effective in controlling unknown or unmanaged risks or losses or in protecting us from governmental investigations or other actions or lawsuits stemming from a failure to comply with such laws or regulations.
If any such actions are instituted against us, and we are not successful in defending ourselves or asserting our rights, those actions could have a material adverse effect on our business, financial condition, results of operations, and prospects. For example, we may be subject to or experience significant civil, criminal, and administrative penalties, damages, monetary fines, individual imprisonment, disgorgement of profits, possible exclusion from participation in Medicare, Medicaid, and other federal healthcare programs, contractual damages, reputational harm, diminished profits and future earnings, additional reporting or oversight obligations if we become subject to a corporate integrity agreement or other agreement to resolve allegations of noncompliance with the law, and curtailment or restructuring of our operations, any of which could adversely affect our ability to operate our business and pursue our strategy.
Current and future legislation may increase the difficulty and cost for us and any future collaborators to obtain marketing approval of and commercialize our product candidates and affect the prices we, or they, may charge for such product candidates.
In the United States and other jurisdictions, there have been, and we expect there will continue to be, a number of legislative and regulatory changes and proposed changes to the healthcare system that could affect our future results of operations. In particular, there have been and continue to be a number of initiatives at the United States. federal and state levels that seek to reduce healthcare costs and improve the quality of healthcare. For example, in March 2010, the Patient Protection and Affordable Care Act, as amended by the Health Care and Education Reconciliation Act (collectively, the ACA), was enacted, which substantially changed the way healthcare is financed by both governmental and private payors. Among the provisions of the ACA of importance to the pharmaceutical and biotechnology industries, which includes biologics, are the following:
|
|
• |
manufacturers and importers of certain branded prescription drugs, including certain biologics, with annual sales of more than $5 million made to or covered by specified federal healthcare programs are required to pay an annual, nondeductible fee according to their market share of all such sales; |
|
|
• |
an increase in the statutory minimum rebates a manufacturer must pay under the Medicaid Drug Rebate Program, to 23.1% of the average manufacturer price for most branded drugs, biologics, and biosimilars, and to 13.0% for generic drugs, and a cap of the total rebate amount for innovator drugs at 100% of the Average Manufacturer Price; |
|
|
• |
a new methodology by which rebates owed by manufacturers under the Medicaid Drug Rebate Program are calculated for certain drugs and biologics that are inhaled, infused, instilled, implanted, or injected, which would include our product candidates; |
|
|
• |
extension of manufacturers’ Medicaid rebate liability to covered drugs dispensed to individuals who are enrolled in Medicaid managed care organizations; |
|
|
• |
expansion of eligibility criteria for Medicaid programs by, among other things, allowing states to offer Medicaid coverage to additional individuals and adding new mandatory eligibility categories for individuals with income at or below 133% of the federal poverty level, thereby potentially increasing manufacturers’ Medicaid rebate liability; |
|
|
• |
expansion of the entities eligible for discounts under the Public Health program, commonly referred to as the “340B Program;” |
|
|
• |
a new requirement to annually report drug samples that manufacturers and distributors provide to physicians, also known as the Physician Payments Sunshine Act; |
|
|
• |
a new Patient-Centered Outcomes Research Institute to oversee, identify priorities in, and conduct comparative clinical effectiveness research, along with funding for such research; |
|
|
• |
establishment of a Center for Medicare Innovation at CMS to test innovative payment and service delivery models to lower Medicare and Medicaid spending, potentially including prescription drug spending; and |
|
|
• |
a licensure framework for follow-on biologic products. |
Since its enactment, there have been judicial, executive and Congressional challenges to certain aspects of the ACA, and we expect there will be additional challenges and amendments to the ACA in the future. On June 17, 2021, the United States Supreme Court dismissed the most recent judicial challenge to the ACA without specifically ruling on the constitutionality of the ACA. Prior to the Supreme Court’s decision, President Biden issued an executive order initiating a special enrollment period from February 15, 2021 through August 15, 2021 for purposes of obtaining health insurance coverage through the ACA marketplace. The executive order also instructed certain governmental agencies to review and reconsider their existing policies and rules that limit access to healthcare.
119
In addition, other legislative changes have been proposed and adopted in the United States since the ACA was enacted, including those which, among other things, have reduced Medicare payments available to several types of providers, including hospitals and cancer treatment centers. These new laws or any other similar laws introduced in the future, as well as regulatory actions that may be taken by CMS, may result in additional reductions in Medicare and other healthcare funding, which could negatively affect target customers for our product candidates and accordingly, our financial operations. Moreover, payment methodologies may be subject to changes as a result of new healthcare legislation and regulatory initiatives. Additionally, individual states in the United States have passed legislation and implemented regulations designed to control pharmaceutical and biological product pricing and costs. Similar developments have occurred outside of the United States, including in the European Union where healthcare budgetary constraints have resulted in restrictions on the pricing and reimbursement of medicines by relevant health service providers. To obtain reimbursement or pricing approval in some European Union member states, we may be required to conduct studies that compare the cost-effectiveness of our product candidates to other therapies that are considered the local standard of care.
It is also possible that additional governmental action is taken in response to address the COVID-19 pandemic. We cannot predict the likelihood, nature, or extent of government regulation that may arise from future legislation or administrative action in the United States or any other jurisdiction. If we or any third parties we may engage or with which we collaborate are slow or unable to adapt to changes in existing requirements or the adoption of new requirements or policies, or if we or such third parties are not able to maintain regulatory compliance, our product candidates may be unable to obtain regulatory approval or lose any regulatory approval that may have been obtained, and we may not achieve or sustain profitability.
Even if we are able to commercialize any product candidate, coverage and adequate reimbursement may not be available or such product candidate may become subject to unfavorable pricing regulations or third-party coverage and reimbursement policies, which would harm our business.
The regulations that govern regulatory approvals, pricing, and reimbursement for drug products vary widely from country to country. Some countries require approval of the sale price of a drug product before it can be marketed. In many countries, the pricing review period begins after marketing approval is granted. In some foreign markets, prescription drug product pricing remains subject to continuing governmental control even after initial approval is granted. As a result, we might obtain regulatory approval for a product in a particular country, but then be subject to price regulations that delay our commercial launch of the product, possibly for lengthy time periods, and negatively impact the revenues we are able to generate from the sale of the product in that country. Adverse pricing limitations may hinder our ability to recoup our investment in one or more product candidates, even if our product candidates obtain regulatory approval.
Our ability to commercialize any products successfully also will depend in part on the extent to which coverage and adequate reimbursement for these products and related treatments will be available from third-party payors, such as government authorities, private health insurers, and other organizations. Even if we succeed in bringing one or more products to the market, these products may not be considered cost-effective, and the amount reimbursed for any products may be insufficient to allow us to sell our products on a competitive basis. Because our product candidates are in the early stages of development, we are unable at this time to determine their cost effectiveness or the likely level or method of coverage and reimbursement. Increasingly, the third-party payors that reimburse patients or healthcare providers are requiring that drug companies provide these payors with predetermined discounts from list prices and are seeking to reduce the prices charged or the amounts reimbursed for drug products. If the price we are able to charge for any products we develop, or the coverage and reimbursement provided for such products, is inadequate in light of our development and other costs, our return on investment could be affected adversely.
There may be significant delays in obtaining reimbursement for newly-approved drug products, and coverage may be more limited than the purposes for which the drug product is approved by the FDA or comparable foreign regulatory authorities. Moreover, eligibility for reimbursement does not imply that any product for which we receive regulatory approval will be reimbursed in all cases or at a rate that covers our costs, including for research, development, manufacture, sale, and distribution.
Interim reimbursement levels for new drug products, if applicable, may also be insufficient to cover our costs and may not be made permanent. Reimbursement rates may be based on payments allowed for lower cost drug products that are already reimbursed, may be incorporated into existing payments for other services, and may reflect budgetary constraints or imperfections in Medicare data. Net prices for drug products may be reduced by mandatory discounts or rebates required by third-party payors and by any future relaxation of laws that presently restrict imports of drug products from countries where they may be sold at lower prices than in the United States. Obtaining coverage and adequate reimbursement for our product candidates may be particularly difficult because of the higher prices often associated with drugs administered under the supervision of a physician. Similarly, because our product candidates are physician-administered injectables, separate reimbursement for the product itself may or may not be available. Instead, the administering physician may or may not be reimbursed for providing the treatment or procedure in which our product is used.
Further, no uniform policy for coverage and reimbursement exists in the United States, and coverage and reimbursement can differ significantly from payor to payor. Third-party payors often follow Medicare coverage policy and payment limitations in setting
120
their own reimbursement rates, but also have their own methods and approval process apart from Medicare determinations. As a result, the coverage determination process is often time-consuming and costly and will likely require us to provide scientific and clinical support for the use of our product candidates to each payor separately, with no assurance that coverage and adequate reimbursement will be applied consistently or obtained in the first instance. Decisions regarding the extent of coverage and amount of reimbursement to be provided for any product candidates that we develop will be made on a payor-by-payor basis. One payor’s determination to provide coverage for a drug does not assure that other payors will also provide coverage and adequate reimbursement for the drug. Additionally, a third-party payor’s decision to provide coverage for a therapy does not imply that an adequate reimbursement rate will be approved.
There have been, and likely will continue to be, legislative and regulatory proposals at the foreign, federal, and state levels directed at broadening the availability of healthcare and containing or lowering the cost of healthcare. We cannot predict what initiatives may be adopted in the future, including repeal or replacement of, or significant revisions to, the Affordable Care Act. The continuing efforts of the government, insurance companies, managed care organizations, and other payors of healthcare services to contain or reduce costs of healthcare or impose price controls may adversely affect:
|
|
• |
the demand for any of our product candidates that may receive regulatory approval; |
|
|
• |
our ability to set a price that we believe is fair for our approved products; |
|
|
• |
our ability to obtain coverage and reimbursement approval for an approved product; |
|
|
• |
our ability to generate revenue and achieve or maintain profitability; |
|
|
• |
the level of taxes that we are required to pay; and |
|
|
• |
the availability of capital. |
Our inability to promptly obtain coverage and adequate reimbursement from third-party payors for the product candidates that we may develop and for which we obtain regulatory approval could have a material and adverse effect on our business, financial condition, results of operations, and prospects.
We face potential liability related to the privacy of personal information, including health information we utilize in the development of products developed from our ex vivo cell engineering platform, as well as information we may obtain from research institutions participating in our clinical trials and directly from individuals.
We and our partners and vendors are subject to various federal, state, and foreign data protection and privacy laws and regulations. If we fail to comply with these laws and regulations, we may be subject to litigation, regulatory investigations, enforcement notices, enforcement actions, fines, and criminal or civil penalties, as well as negative publicity, reputational harm, and a potential loss of business.
In the United States, our and our partners’ operations are subject to numerous federal and state laws and regulations, including state data breach notification laws and federal and state data privacy laws and regulations that govern the collection, use, disclosure, and protection of health information and other personal information. For example, most healthcare providers, including research institutions from which we obtain patient health information, are subject to data privacy and security regulations promulgated under HIPAA, as amended by the Health Information Technology for Economic and Clinical Health Act of 2009 (HITECH). Depending on the facts and circumstances, we could be subject to significant penalties if we violate HIPAA. For example, under HIPAA, we could potentially face substantial criminal or civil penalties if we knowingly receive protected health information from a HIPAA-covered healthcare provider or research institution that has not satisfied HIPAA’s requirements for disclosure of such health information, or otherwise violate applicable HIPAA requirements related to the protection of such information. Even when HIPAA does not apply, failing to take appropriate steps to keep consumers’ personal information secure may constitute a violation of the Federal Trade Commission Act.
Certain of the research materials we use in our therapeutic research and development efforts, as well as stem cell lines used as starting material in our ex vivo cell engineering product candidates, are derived from human sources, which may contain sensitive identifiable personal information regarding the donor. In addition, once we commence clinical trials, we or our partners may maintain or otherwise have access to sensitive identifiable personal information, including health information, that we receive throughout the clinical trial process, in the course of our research collaborations, and directly from individuals (or their healthcare providers) who enroll in our patient assistance programs. We may become subject to further obligations under HIPAA as a result of our access to such information. In addition, our collection of personal information generally, including information of our employees or future patients, may subject us to state data privacy laws governing the processing of personal information and requiring notification of affected individuals and state regulators in the event of a data breach involving such personal information. These state laws include the California Consumer Privacy Act (CCPA) and its related regulations, and, once effective, the recently approved California Privacy
121
Rights Act (CPRA) amending the CCPA, which establish data privacy rights for residents of the State of California, with corresponding obligations on businesses related to transparency, deletion, and opt-out of the selling of personal information, and grant a private right of action for individuals in the event of certain security breaches.
California voters approved the CPRA in the November 3, 2020 election. Effective January 1, 2023, the CPRA will significantly modify the CCPA, including by expanding consumers’ rights with respect to certain sensitive personal information. The CPRA also creates a new state agency that will be vested with authority to implement and enforce the CCPA and the CPRA. New legislation relating to data privacy and security has been proposed or enacted in various other states and at the federal level. Such legislation will continue to shape the data privacy environment nationally. Certain state laws may be more stringent or broader in scope, or offer greater individual rights, with respect to confidential, sensitive, and personal information than federal, international or other state laws, and such laws may differ from each other and have potentially conflicting requirements that would make compliance challenging, require us to expend significant resources achieve compliance, and restrict our ability to process certain personal information.
Any clinical trial programs and research collaborations that we engage in outside the United States may implicate international data protection laws, including, in Europe, the General Data Protection Regulation (GDPR). The GDPR imposes stringent operational requirements for data controllers and data processors of personal data. Among other things, the GDPR requires that detailed notices be provided to clinical trial subjects and investigators, as well as maintenance of certain security levels for personal data and notification of data processing obligations or security incidents to appropriate data protection authorities or data subjects. Further, following the United Kingdom’s withdrawal from the European Union, effective as of December 31, 2020, we will be required to comply with both the GDPR and the GDPR as incorporated into United Kingdom national law with respect to any clinical trial data generated from the European Union and the United Kingdom, respectively, which may have differing requirements.
One particularly sensitive issue under these European Union data privacy laws involves the transfer of personal data from the European Economic Area (EEA) to other jurisdictions. Recent legal developments in Europe have created complexity and uncertainty regarding the legality of and requirements with respect to transfers of personal data from the EEA to the United States and other countries in which we or our partners or service providers may operate. For example, on July 16, 2020, the Court of Justice of the European Union (CJEU) invalidated the EU-US Privacy Shield Framework (Privacy Shield), under which personal data could previously be transferred from the EEA to United States entities that had self-certified under the Privacy Shield scheme. The CJEU decision also created additional obligations and uncertainty regarding the use standard contractual clauses for such data transfers. As government authorities issue further guidance on personal data export mechanisms or start aggressively taking enforcement action based on such guidance or the CJEU decision, we could be subject to additional costs, complaints, regulatory investigations or fines. If we are unable to transfer personal data between and among countries and regions in which we or our partners or service providers operate, it could adversely affect the manner in which we operate our business, affect the geographical location or segregation of our relevant systems and operations, and adversely affect our financial results. These laws and regulations may also apply to vendors that store or otherwise process personal data on our behalf, such as information technology or other vendors. If our data privacy or security measures fail to comply with applicable data privacy laws, or if a vendor misuses data we have provided to it or fails to safeguard such data, or otherwise fails to comply with such laws, we may be subject to litigation, regulatory investigations, enforcement notices, or enforcement actions imposing fines or requiring us to change the way we use personal data, as well as negative publicity, reputational harm, and a potential loss of business.
We expect that we will need to expend significant capital and other resources to ensure ongoing compliance with applicable data privacy and security laws. Claims that we have violated individuals’ privacy rights or breached our contractual obligations related to data privacy and security, even if we are not found liable, could be expensive and time-consuming to defend and could result in negative publicity that could harm our business. Moreover, even if we take all necessary action to comply with legal and regulatory requirements, we could be subject to a data breach or other unauthorized access of personal information, which could subject us to fines and penalties, as well as litigation and reputational damage.
If we fail to keep apprised of and comply with applicable international, federal, state, or local regulatory requirements and changes thereto, we could be subject to a range of regulatory actions that could affect our or any collaborators’ ability to seek to commercialize our product candidates. Any threatened or actual government enforcement action, or litigation where private rights of action are available, could also generate negative publicity, damage our reputation, result in liabilities, fines, and loss of business, and require that we devote substantial resources that could otherwise be used in support of other aspects of our business.
122
Risks Related to Our Limited Operating History, Financial Condition and Need for Additional Capital
We are a preclinical-stage biotechnology company and have incurred significant losses since our inception, and we expect to incur losses for the foreseeable future. We have no products approved for commercial sale and may never achieve or maintain profitability.
We are a preclinical-stage biotechnology company with a limited operating history. Biotechnology product development is a highly speculative undertaking and involves a substantial degree of risk. We have incurred significant losses since inception, have not generated any revenue from product sales, and have financed our operations historically through private placements of our convertible preferred stock and, more recently, through our IPO. We expect that it will be several years, if ever, before we have a commercialized product and generate revenue from product sales. We had net losses of $355.9 million, $285.3 million and $130.8 million for the year ended December 31, 2021, 2020, and 2019, respectively. As of December 31, 2021, we had an accumulated deficit of $785.4 million. Our losses have resulted principally from expenses incurred for the research and development of our ex vivo and in vivo cell engineering platforms, management and administrative costs, and other expenses incurred while building our business infrastructure.
We expect our operating losses and expenses will continue to increase substantially for the foreseeable future, including as we:
|
|
• |
expand our research and development efforts; |
|
|
• |
advance and expand the capabilities of our ex vivo and in vivo cell engineering platforms; |
|
|
• |
identify additional product candidates; |
|
|
• |
advance preclinical development of our current product candidates and initiate additional preclinical studies, including with respect to future product candidates; |
|
|
• |
commence and advance through clinical studies of our current and future product candidates; |
|
|
• |
establish our manufacturing capability, including developing our contract development and manufacturing organization (CDMO) relationships and building our internal manufacturing facilities; |
|
|
• |
acquire and license technologies aligned with our ex vivo and in vivo cell engineering platforms; |
|
|
• |
seek regulatory approval of our current and future product candidates; |
|
|
• |
engage in commercialization activities, including product manufacturing, marketing, sales, and distribution for any of our product candidates for which we obtain marketing approval; |
|
|
• |
expand our operational, financial, and management systems and increase personnel, including those required to support our preclinical and clinical development, manufacturing, and potential future commercialization efforts; |
|
|
• |
continue to develop, prosecute, and defend our intellectual property portfolio; and |
|
|
• |
incur additional legal, accounting, and other expenses necessary to operate our business, including the costs associated with operating as a public company. |
We have devoted a significant portion of our financial resources and efforts to building our organization, developing our ex vivo and in vivo cell engineering platforms, identifying and developing potential product candidates, executing preclinical studies, establishing manufacturing capabilities, acquiring technology, organizing and staffing the company, developing and executing our business plan, establishing our intellectual property portfolio, raising capital, and providing general and administrative support for these operations. We are in the early stages of development of our product candidates, have not yet commenced any clinical trials for any of our product candidates, and have not completed development or commercialization of any product candidate.
To become and remain profitable, we must succeed in identifying, developing, obtaining regulatory approval for, and eventually commercializing products that generate significant revenue. This will require us to be successful in a range of challenging activities, including completing preclinical studies and clinical trials of our product candidates, continuing to discover and develop additional product candidates, obtaining regulatory approval for any product candidates that successfully complete clinical trials, accessing manufacturing capacity, establishing marketing capabilities, and commercializing and ultimately selling any products for which we may obtain regulatory approval. We may never succeed in any or all of these activities and, even if we do, we may never generate revenue that is sufficient to achieve profitability. Even if we do achieve profitability, we may not be able to sustain profitability or meet outside expectations for our profitability. If we are unable to achieve or sustain profitability or to meet outside expectations for our profitability, the value of our shares of common stock could be materially adversely affected.
Because of the numerous risks and uncertainties associated with biopharmaceutical product development, we are unable to accurately predict the timing or increases in the amount of expenses we will incur or when, or if, we will be able to achieve profitability. If we are required by the FDA or comparable foreign regulatory authorities to perform studies in addition to those we
123
currently anticipate, or if there are any delays in commencing or completing our clinical trials or the development of any of our product candidates, our expenses could increase and our ability to obtain commercial revenue could be further delayed and become more uncertain, which will have a material adverse impact on our business.
We will require additional funding in order to finance our operations. If we are unable to raise capital when needed, or on acceptable terms, we could be forced to delay, reduce, or eliminate our product development programs or commercialization efforts.
Developing biopharmaceutical products, including conducting preclinical studies and clinical trials, is a very time-consuming, expensive, and uncertain process that takes years to complete. As described above, our operations have consumed substantial amounts of cash since inception, and we expect our expenses to increase in connection with our ongoing activities. Accordingly, we will need to obtain substantial additional funding in connection with our continuing operations. If we are unable to raise capital when needed or on attractive terms, we could be forced to delay, reduce, or eliminate our research and development programs or any future commercialization efforts.
As of December 31, 2021, we had $746.9 million in cash, cash equivalents, and marketable securities. Based on our current business plans, we believe that our existing cash, cash equivalents, and marketable securities as of December 31, 2021 will be sufficient to fund our operating expenses and capital expenditure requirements for at least the next 12 months. We have based this estimate on assumptions that may prove to be wrong, and we could use our capital resources more quickly than we currently expect, which could require us to seek additional funds sooner than planned, including through public or private equity or debt financings or other sources, such as strategic collaborations. In addition, we may seek additional capital due to favorable market conditions or strategic considerations even if we believe we have sufficient funds for our current or future operating plans. Attempting to secure additional financing may divert our management from our day-to-day activities, which may adversely affect our ability to develop our product candidates. Our future capital requirements will depend on many factors, including:
|
|
• |
the scope, timing, progress, costs, and results of discovery, preclinical development, and clinical trials for our current or future product candidates; |
|
|
• |
the number and scope of clinical trials required for regulatory approval of our current or future product candidates; |
|
|
• |
the costs, timing, and outcome of regulatory review of our current or future product candidates; |
|
|
• |
the cost associated with building our manufacturing capabilities, as well as costs associated with the manufacturing of clinical and commercial supplies of our current or future product candidates; |
|
|
• |
the costs and timing of future commercialization activities, including manufacturing, marketing, sales, and distribution, for any of our product candidates for which we receive marketing approval; |
|
|
• |
the costs and timing of preparing, filing, and prosecuting patent applications, maintaining and enforcing our intellectual property rights, and defending any intellectual property-related claims, including any claims by third parties that we are infringing upon their intellectual property rights; |
|
|
• |
our ability to maintain existing, and establish new, strategic collaborations, licensing, or other arrangements and the financial terms of any such agreements, including the timing and amount of any future milestone, royalty, or other payments due under any such agreement; |
|
|
• |
the revenue, if any, received from commercial sales of our product candidates for which we receive marketing approval; |
|
|
• |
the expenses required to attract, hire, and retain skilled personnel; |
|
|
• |
the costs of operating as a public company; |
|
|
• |
our ability to establish a commercially viable pricing structure and obtain approval for coverage and adequate reimbursement from third-party and government payors; |
|
|
• |
potential interruptions or delays resulting from factors related to the ongoing COVID-19 pandemic; |
|
|
• |
the effect of competing technological and market developments; and |
|
|
• |
the extent to which we acquire or invest in businesses, products, and technologies. |
Our ability to raise additional funds will depend on financial, economic, political, and market conditions and other factors over which we may have no or limited control. Market volatility resulting from the ongoing COVID-19 pandemic or other factors, such as the recent escalation in conflict between Russia and Ukraine, could also adversely impact our ability to access capital as and when
124
needed. Additional funds may not be available when we need them, on terms and at a cost that are acceptable to us, or at all. If adequate funds are not available to us on a timely basis, we could be required to:
|
|
• |
delay, limit, reduce, or terminate preclinical studies, clinical trials, or other research and development activities, or eliminate one or more of our development programs altogether; or |
|
|
• |
delay, limit, reduce, or terminate our efforts to access manufacturing capacity or establish and operationalize our manufacturing facility, establish sales and marketing capabilities, or other activities that may be necessary to commercialize any product candidates for which we obtain regulatory approval, or reduce our flexibility in developing or maintaining our sales and marketing strategy with respect to any product candidates for which we obtain regulatory approval. |
Raising additional capital may cause dilution to our stockholders, restrict our operations, or require us to relinquish rights to our technologies or product candidates.
Until such time, if ever, as we can generate substantial product revenue, we expect to finance our operations with our existing cash, cash equivalents, and marketable securities, any future equity or debt financings, and upfront, milestone, and royalty payments received under any future licenses or collaborations. If we raise additional capital through the sale of equity or debt securities, existing stockholders’ ownership interests will be diluted, and the terms of these securities may include liquidation or other preferences that adversely affect the rights of our stockholders. In addition, the possibility of such issuance may cause the market price of our common stock to decline. Debt financing, if available, may result in increased fixed payment obligations and involve agreements that include covenants limiting or restricting our ability to take specific actions, such as incurring additional debt, making capital expenditures, declaring dividends, or acquiring, selling, or licensing intellectual property rights or assets, which could adversely impact our ability to conduct our business.
If we raise additional funds through collaborations, strategic alliances, or marketing, distribution, or licensing arrangements with third parties, we may have to relinquish valuable rights to our intellectual property, technologies, future revenue streams, or product candidates or grant licenses on terms that may not be favorable to us. We could also be required to seek funds through arrangements with collaborators or others at an earlier stage than otherwise would be desirable. Any of these occurrences may have a material adverse effect on our business, operating results, and prospects.
Our success payment and contingent consideration obligations in our license and acquisition agreements may result in dilution to our stockholders, drain our cash resources, or cause us to incur debt to satisfy the payment obligations.
We agreed to make success payments, payable in cash, pursuant to our license agreement with the President and Fellows of Harvard College (Harvard) and contingent consideration and success payments, payable in cash or stock, pursuant to our acquisition agreement with Cobalt Biomedicine, Inc. (Cobalt). The success payments to Harvard (Harvard Success Payments) are based on increases in the fair value of our common stock. The potential Harvard Success Payments are based on multiples of increased value ranging from 5x to 40x based on a comparison of the per share fair value of our common stock relative to the original $4.00 issuance price at pre-determined valuation measurement dates. The amount of the Harvard Success Payments will not exceed an aggregate of $175.0 million, which maximum amount would only be payable upon a 40x increase in the fair value of our common stock. The Harvard Success Payments can be achieved over a maximum of 12 years from the effective date of the agreement. The valuation measurement dates for the Harvard Success Payments are triggered by events that include the one-year anniversary of our IPO, and periodically thereafter, the date of the consummation of a merger, an asset sale, or the sale of the majority of the shares held by our Series A convertible preferred stockholders, and the last day of the term of the Harvard Success Payments. If a higher success payment tier is met at the same time a lower tier is first met, both tiers will be owed. Any previous success payments made to Harvard are credited against the success payment owed as of any valuation measurement date so that Harvard does not receive multiple success payments in connection with the same threshold. The first valuation measurement date occurred in February 2022 following the one-year anniversary of our IPO. The threshold was not met, and therefore no payment was made as of measurement date.
In connection with the Cobalt acquisition, we are obligated to pay contingent consideration (Cobalt Contingent Consideration) of up to an aggregate of $500.0 million to certain former Cobalt stockholders upon our achievement of certain pre-defined development milestones. Additionally, we are obligated to pay a success payment to certain Cobalt stockholders (Cobalt Success Payment) of $500.0 million if, at pre-determined valuation measurement dates, including the closing of our IPO and periodically thereafter, our market capitalization equals or exceeds $8.1 billion, and we are advancing a program based on the fusogen technology in a clinical trial pursuant to an IND, or have filed for, or received approval for, a biologics license application (BLA) or new drug application (NDA) with respect to a program based on the fusogen technology. In addition to our IPO, a valuation measurement date would be triggered upon a change of control if at least one of our programs based on the fusogen technology is the subject of an active research program at the time of such change of control. If there is a change of control and our market capitalization is below $8.1 billion as of the date of the change of control, the amount of the potential Cobalt Success Payment will decrease, and the amount of potential Cobalt Contingent Consideration will increase. The term of the Cobalt Success Payment is 20 years from the date of the Cobalt acquisition. See Note 3, Acquisitions, to our consolidated financial statements included elsewhere in this Annual Report for
125
details on the amount of the potential Cobalt Success Payment and potential Cobalt Contingent Consideration if there is a change of control based on various thresholds for our market capitalization on such change of control date.
In order to satisfy our obligations to make these success payments, if and when they are triggered, we may issue equity or convertible debt securities that may cause dilution to our stockholders, or we may use our existing cash or incur debt to satisfy the success payment obligations in cash, which may adversely affect our financial position. In addition, these success payments may impede our ability to raise money in future public offerings of debt or equity securities or to obtain a third-party line of credit. The first valuation measurement date for the Harvard Success Payments occurred following the one-year anniversary of our IPO, but it did not trigger a Harvard Success Payment. See Note 4, License and collaboration agreements, to our consolidated financial statements included elsewhere in this Annual Report for the per share common stock prices that would trigger a Harvard Success Payment and the corresponding payment amount. The first valuation measurement date for the Cobalt Success Payment was our IPO, but our IPO did not trigger any such payment. However, the triggering of such Cobalt Success Payment depends on both our progress on fusogen-related product candidates as well as our market capitalization, which is unpredictable and may fluctuate significantly from quarter to quarter and year to year.
The contingent consideration and success payment obligations in our license and acquisition agreements may cause our operating results, net losses, and financial condition as reported by United States generally accepted accounting principles to fluctuate significantly from quarter to quarter and year to year, which may reduce the usefulness of our financial statements.
Our success payment and contingent consideration obligations under our license and acquisition agreements are recorded as liabilities on our consolidated balance sheets. Under United States generally accepted accounting principles (GAAP), we are required to estimate the fair value of these liabilities as of each quarter end, with changes in the estimated fair value recorded in research and development-related success payments and contingent consideration. Factors that may lead to increases or decreases in the estimated fair value of the success payment liabilities include, among others, changes in the value of our common stock and market capitalization, changes in volatility, the estimated number and timing of valuation measurement dates, the term of the success payments, and changes in the risk-free interest rate. Factors that may lead to increases or decreases in the estimated fair value of our contingent consideration obligations include, among others, the estimated likelihood and timing within which milestones may be achieved and the estimated discount rates. A small change in the inputs and related assumptions with respect to our success payment and contingent consideration liabilities may result in a relatively large change in the estimated valuation and associated liabilities and resulting expense or gain. As a result, our operating results, net losses, and financial condition as reported by GAAP may fluctuate significantly from quarter to quarter and year to year for reasons unrelated to our operations, which may reduce the usefulness of our GAAP financial statements. For example, as of December 31, 2021, September 30, 2021, June 30, 2021, and March 31, 2021, the estimated aggregate fair value of the Cobalt Success Payment and Harvard Success Payments liabilities was $102.5 million, $134.2 million, $109.0 million, and $192.2 million, respectively, and the estimated fair value of the Cobalt Contingent Consideration was $153.7 million, $132.0 million, $140.5 million, and $133.3 million, respectively.
For the three and twelve months ended December 31, 2021, we recorded expenses of $21.8 million and $31.8 million, respectively, related to the change in the estimated fair value of the Cobalt Contingent Consideration. For the three and twelve months ended December 31, 2021, we recorded a gain of $31.7 million and expense of $26.0 million, respectively, related to the aggregate change in the estimated fair value of these success payment liabilities. For the Harvard Success Payments, keeping all other variables constant, a hypothetical 20% increase in our common stock price at December 31, 2021 from $15.48 per share to $18.58 per share would have decreased the gain recorded in the three months ended December 31, 2021 associated with the success payment liability by $4.0 million to $4.4 million. A hypothetical 20% decrease in the common stock price from $15.48 per share to $12.38 per share would have increased the gain recorded in three months ended December 31, 2021 by $3.7 million to $12.1 million. For the Cobalt Success Payment, keeping all other variables constant, a hypothetical 20% increase in our market capitalization at December 31, 2021 from $2.9 billion to $3.5 billion would have decreased the gain recorded in the three months ended December 31, 2021 associated with the success payment liability by $20.8 million to $2.5 million. A hypothetical 20% decrease in our market capitalization from $2.9 billion to $2.3 billion would have increased the gain recorded in the three months ended December 31, 2021 by $21.3 million to $44.5 million.
We have incurred net losses since our inception and expect to continue to incur net losses for the foreseeable future. It is possible that future fluctuations in the price of our common stock and market capitalization and the resulting change in the estimated fair value of our success payment liabilities could lead us to record net income in a future period despite us incurring operating losses and negative cash flows during such period. Alternatively, significant stock appreciation during a future period could lead to a significant increase in our recorded GAAP net loss.
Our limited operating history may make it difficult to evaluate our prospects and likelihood of success.
We are a preclinical-stage biopharmaceutical company with a limited operating history upon which to evaluate our business and prospects. Since our inception in July 2018, we have devoted substantially all of our resources and efforts to building our organization,
126
developing our ex vivo and in vivo cell engineering platforms, identifying and developing potential product candidates, executing preclinical studies, establishing manufacturing capability, acquiring technology, organizing and staffing the company, developing and executing our business plan, establishing and securing our intellectual property portfolio, raising capital, and providing general and administrative support for these operations. Since all of our product candidates are still in preclinical development, we have not yet demonstrated our ability to successfully commence or complete any clinical trials, including Phase 3 or other pivotal clinical trials, obtain regulatory approvals, manufacture a commercial-scale product or arrange for a third party to do so on our behalf, or conduct sales and marketing activities necessary for successful product commercialization. Additionally, we expect our financial condition and operating results to continue to fluctuate significantly from period to period due to a variety of factors, many of which are beyond our control. Consequently, predictions about our future success or viability are difficult to make and may not be as accurate as they could be if we had a longer operating history.
Risks Related to Commercialization of Our Product Candidates
We operate in highly competitive and rapidly changing industries, which may result in others discovering, developing, or commercializing competing products before or more successfully than we do.
The biotechnology and pharmaceutical industries are highly competitive and subject to significant and rapid technological change. Our success is highly dependent on our ability to discover, develop, and obtain marketing approval for new and innovative products on a cost-effective basis and to market them successfully. In doing so, we face and will continue to face intense competition from a variety of businesses, including large pharmaceutical companies, biotechnology companies, academic institutions, government agencies, and other public and private research organizations. These organizations may have significantly greater resources than we do and conduct similar research, seek patent protection, and establish collaborative arrangements for research, development, manufacturing, and marketing of products that compete with our product candidates. Mergers and acquisitions in the pharmaceutical and biotechnology industries may result in even more resources being concentrated in our competitors. Competition may increase further as a result of advances in the commercial applicability of technologies and greater availability of capital for investment in these industries.
With the proliferation of new drugs and therapies for our target indications, and as new technologies become available, we expect to face increasingly intense competition. If we fail to stay at the forefront of technological change, we may be unable to compete effectively. Any product candidates that we successfully develop and commercialize will compete with existing therapies and new therapies that may become available in the future. The highly competitive nature of and rapid technological changes in the biotechnology and pharmaceutical industries could render our product candidates or our technologies obsolete, less competitive, or uneconomical. Our competitors may, among other things:
|
|
• |
have significantly greater financial, manufacturing, marketing, drug development, technical, and human resources than we do; |
|
|
• |
develop and commercialize products that are safer, more effective, less expensive, more convenient or easier to administer, or have fewer or less severe side effects; |
|
|
• |
obtain quicker regulatory approval; |
|
|
• |
establish proprietary positions covering our products and technologies; |
|
|
• |
implement more effective approaches to sales and marketing; or |
|
|
• |
form more advantageous strategic alliances. |
Should any of these factors occur, our business, financial condition, and results of operations could be materially adversely affected.
In addition, our collaborators may decide to market and sell products that compete with the product candidates that we have agreed to license to them, which could have a material adverse effect on our future business, financial condition, and results of operations.
Smaller and other early-stage companies may also prove to be significant competitors, particularly through collaborative arrangements with large and established companies. We currently and in the future will compete with these third parties in recruiting and retaining qualified scientific and management personnel, establishing clinical trial sites, and recruiting patients for clinical trials, as well as in acquiring technologies complementary to, or necessary for, our programs. See the subsection titled “Business—Competition” in this Annual Report.
127
Market opportunity and market growth for our product candidates may prove to be smaller than we initially estimated, and even if the markets in which we compete achieve the forecasted growth, our business may not grow at similar rates, or at all.
We intend to initially focus our product candidate development on treatments for various diseases caused by missing or damaged cells. Our projections of addressable patient populations within any particular disease state that may benefit from treatment with our product candidates are based on our estimates. Market opportunity estimates and growth forecasts are subject to significant uncertainty and are based on assumptions and estimates. These estimates, which have been derived from a variety of sources, including scientific literature, surveys of clinics, patient foundations, and market research, may prove to be incorrect. Further, new studies may change the estimated incidence or prevalence of these diseases. Additionally, the potentially addressable patient population for our product candidates may not ultimately be amenable to treatment with our product candidates. Our market opportunity may also be limited by future competitor therapies that enter the market. If any of our estimates proves to be inaccurate, the market opportunity for any product candidate that we or our strategic partners develop could be significantly diminished, which would have an adverse material impact on our business.
In particular, certain of our product candidates are intended to address cancer, and, in particular, B cell malignancies. Cancer therapies are sometimes characterized as first line, second line, or third line and beyond, and the FDA often approves new therapies initially only for a particular line of use. When cancer is detected early enough, first line therapy is sometimes adequate to cure the cancer or prolong life without a cure. Whenever first line therapy, which usually consists of chemotherapy, antibody drugs, tumor-targeted small molecules, hormone therapy, radiation therapy, surgery, or a combination of these, proves unsuccessful, second line therapy may be administered. Second line therapies often consist of more chemotherapy, radiation, antibody drugs, tumor-targeted small molecules, or a combination of these. Third line therapies can include chemotherapy, antibody drugs, and small molecule tumor-targeted therapies, more invasive forms of surgery, and new technologies. The use of certain classes of therapies, including CAR T therapies, has been limited to a subset of patients with relapsed or refractory disease. Our projections of both the number of people who have the cancers we are targeting, as well as the subset of people with these cancers who are in a position to receive a particular line of therapy and who have the potential to benefit from treatment with our product candidates, are based on our beliefs and estimates. Consequently, even if our product candidates are approved for a later line of therapy, the number of patients that may be eligible for treatment with our product candidates may turn out to be much lower than expected.
We currently have no marketing, sales, or distribution infrastructure and we intend to either establish a sales and marketing infrastructure or outsource this function to a third party. Each of these commercialization strategies carries substantial risks to us.
We currently have no marketing, sales, or distribution capabilities because all of our product candidates are still in preclinical development. If one or more of our product candidates complete clinical development and receive regulatory approval, we intend to either establish a sales and marketing organization with technical expertise and supporting distribution capabilities to commercialize our product candidates in a legally compliant manner, or to outsource this function to a third party. There are risks involved if we decide to establish our own sales and marketing capabilities or enter into arrangements with third parties to perform these functions. To the extent that we enter into collaboration agreements with respect to marketing, sales, or distribution, our product revenue may be lower than if we directly marketed or sold any approved products. Such collaborative arrangements with partners may place the commercialization of our products outside of our control and would subject us to a number of risks, including that we may not be able to control the amount or timing of resources that our collaborative partner devotes to our products or that our collaborator’s willingness or ability to complete its obligations, and our ability to complete our obligations under these arrangements, may be adversely affected by business combinations or significant changes in our collaborator’s business strategy.
If we are unable to enter into these arrangements on acceptable terms, or at all, we may not be able to successfully commercialize any products for which we receive regulatory approval. If we are not successful in commercializing any approved products, either on our own or through collaborations with one or more third parties, our ability to generate product revenue will suffer and we may incur significant additional losses, which would have a material adverse effect on our business, financial condition, and results of operations.
Our product candidates for which we intend to seek approval as biologic products may face competition sooner than anticipated.
The ACA includes a subtitle called the Biologics Price Competition and Innovation Act of 2009 (BPCIA), which created an abbreviated approval pathway for biological products that are biosimilar to or interchangeable with an FDA-licensed reference biological product. Under the BPCIA, an application for a highly similar or “biosimilar” product may not be submitted to the FDA until four years following the date that the reference product was first approved by the FDA. In addition, the approval of a biosimilar product may not be made effective by the FDA until 12 years from the date on which the reference product was first approved. During this 12-year period of exclusivity, another company may still market a competing version of the reference product if the FDA approves a full BLA for the competing product containing the sponsor’s own preclinical data and data from adequate and well-controlled clinical trials to demonstrate the safety, purity, and potency of the competing product. The law is complex and is still being interpreted and implemented by the FDA. As a result, its ultimate impact, implementation, and meaning are subject to uncertainty. In addition, complexities associated with the larger, and often more complex, structures of biological products, such as cell and gene
128
products that we are developing, as well as the processes by which such products are manufactured, pose significant hurdles to implementation of the abbreviated approval pathway that are still being worked out by the FDA.
We believe that any of our product candidates that may be approved as a biological product under a BLA should qualify for the 12-year period of exclusivity. However, there is a risk that this exclusivity could be shortened due to congressional action or otherwise, or that the FDA will not consider our product candidates to be reference products for competing products, potentially creating the opportunity for competition sooner than anticipated. Other aspects of the BPCIA, some of which may impact the BPCIA exclusivity provisions, have also been the subject of recent litigation. Moreover, the extent to which a biosimilar, once approved, will be substituted for any one of our reference products in a way that is similar to traditional generic substitution for non-biological products is not yet clear, and will depend on a number of marketplace and regulatory factors that are still developing.
Jurisdictions outside the United States have also established abbreviated pathways for regulatory approval of biological products that are biosimilar to earlier approved reference products. For example, the European Union has had an established regulatory pathway for biosimilars since 2004. However, biosimilars can only be authorized once the period of data exclusivity on the reference biological medicine has expired.
The increased likelihood of biosimilar competition has increased the risk of loss of innovators’ market exclusivity. Due to this risk, and uncertainties regarding patent protection, we are not currently able to predict with certainty the length of market exclusivity for any particular product candidate that may receive marketing approval based solely on the expiration of the relevant patent(s) or the current forms of regulatory exclusivity. There may also be future changes in United States regulatory law that might reduce biological product regulatory exclusivity. The loss of market exclusivity for any product for which we receive regulatory approval could materially and negatively affect our ability to generate revenues, which could prevent us from generating adequate or sufficient revenues and being able to achieve or sustain profitability.
Risks Related to Ownership of Our Common Stock
Our principal stockholders and management own a significant percentage of our stock and will be able to exert significant control over matters subject to stockholder approval.
As of December 31, 2021, our executive officers, directors, holders of 5% or more of our capital stock, and their respective affiliates, owned approximately 64.7% of our outstanding voting stock. Therefore, these stockholders have the ability to influence us through this ownership position. These stockholders may be able to determine all matters requiring stockholder approval. For example, these stockholders may be able to control elections of directors, amendments of our organizational documents, or approval of any merger, sale of assets, or other major corporate transaction. This may prevent or discourage unsolicited acquisition proposals or offers for our common stock that other stockholders may feel are in their best interests.
Future sales of our common stock in the public market could cause our common stock price to fall.
Our common stock price could decline as a result of sales of a large number of shares of common stock or the perception that these sales could occur. These sales, or the possibility that these sales may occur, might also make it more difficult for us to sell equity securities in the future at a time and price that we deem appropriate. As of December 31, 2021, 189.3 million shares of our common stock were outstanding. Substantially all shares of common stock sold in our IPO (excluding any shares sold to our directors or officers in the directed share program) are freely tradable without restriction or further registration under the Securities Act of 1933, as amended (Securities Act), unless held by our “affiliates” as defined in Rule 144 under the Securities Act. Shares issued upon the exercise of stock options outstanding under our equity incentive plans or pursuant to future awards granted under those plans will become available for sale in the public market to the extent permitted by the provisions of applicable vesting schedules, as well as Rules 144 and 701 under the Securities Act. We also register the offer and sale of all shares of common stock that we may issue under our equity compensation plans. Accordingly, these shares may be sold in the public market upon issuance. In addition, in the future, we may issue additional shares of common stock, or other equity or debt securities convertible into common stock, in connection with a financing, acquisition, employee arrangement, or otherwise. Any such issuance could result in substantial dilution to our existing stockholders and could cause the price of our common stock to decline.
We do not currently intend to pay dividends on our common stock and, consequently, our stockholders’ ability to achieve a return on their investment will depend on appreciation of the value of our common stock.
We have never declared or paid cash dividends on our common stock. We currently intend to retain all available funds and any future earnings to support operations and to finance the growth and development of our business. We do not intend to declare or pay any cash dividends on our capital stock in the foreseeable future. As a result, any investment return on our common stock will depend upon increases in the value for our common stock, which is not certain.
129
Provisions in our amended and restated certificate of incorporation and our amended and restated bylaws and Delaware law might discourage, delay, or prevent a change in control of our company or changes in our management and, therefore, depress the market price of our common stock.
Our amended and restated certificate of incorporation and amended and restated bylaws contain provisions that could depress the market price of our common stock by acting to discourage, delay, or prevent a change in control of our company or changes in our management that the stockholders of our company may deem advantageous. These provisions, among other things:
|
|
• |
establish a staggered Board divided into three classes serving staggered three-year terms, such that not all members of the Board are elected at one time; |
|
|
• |
authorize our Board to issue new series of preferred stock without stockholder approval and create, subject to applicable law, a series of preferred stock with preferential rights to dividends or our assets upon liquidation, or with superior voting rights to our existing common stock; |
|
|
• |
eliminate the ability of our stockholders to call special meetings of stockholders; |
|
|
• |
eliminate the ability of our stockholders to fill vacancies on our Board; |
|
|
• |
establish advance notice requirements for nominations for election to our Board or for proposing matters that can be acted upon by stockholders at our annual stockholder meetings; |
|
|
• |
permit our Board to establish the number of directors; |
|
|
• |
provide that our Board is expressly authorized to make, alter, or repeal our bylaws; |
|
|
• |
provide that stockholders can remove directors only for cause and only upon the approval of not less than 66 2/3% of all outstanding shares of our voting stock; |
|
|
• |
require the approval of not less than 66 2/3% of all outstanding shares of our voting stock to amend our bylaws and specific provisions of our certificate of incorporation; and |
|
|
• |
limit the jurisdictions in which certain stockholder litigation may be brought. |
As a Delaware corporation, we will be subject to the anti-takeover provisions of Section 203 of the Delaware General Corporation Law, which prohibits a Delaware corporation from engaging in a business combination specified in the statute with an interested stockholder (as defined in the statute) for a period of three years after the date of the transaction in which the person first becomes an interested stockholder, unless the business combination is approved in advance by a majority of the independent directors or by the holders of at least two-thirds of the outstanding disinterested shares. The application of Section 203 of the Delaware General Corporation Law could also have the effect of delaying or preventing a change of control of our company.
Our certificate of incorporation provides that the Court of Chancery of the State of Delaware will be the exclusive forum for substantially all disputes between us and our stockholders, which could limit our stockholders’ ability to obtain a favorable judicial forum for disputes with us or our directors, officers, or employees.
Our amended and restated certificate of incorporation provides that, unless we consent in writing to the selection of an alternative forum, the sole and exclusive forum, to the fullest extent permitted by law, for (i) any derivative action or proceeding brought on our behalf, (ii) any action asserting a breach of a fiduciary duty owed by any director, officer, or other employee to us or our stockholders, (iii) any action asserting a claim against us or any director, officer, or other employee arising pursuant to the Delaware General Corporation Law, (iv) any action to interpret, apply, enforce, or determine the validity of our second amended and restated certificate of incorporation or amended and restated bylaws, or (v) any other action asserting a claim that is governed by the internal affairs doctrine, shall be the Court of Chancery of the State of Delaware (or another state court or the federal court located within the State of Delaware if the Court of Chancery does not have or declines to accept jurisdiction), in all cases subject to the court’s having jurisdiction over indispensable parties named as defendants. In addition, our amended and restated certificate of incorporation provides that the federal district courts of the United States will be the exclusive forum for resolving any complaint asserting a cause of action arising under the Securities Act, but that the forum selection provision will not apply to claims brought to enforce a duty or liability created by the Securities Exchange Act of 1934, as amended (Exchange Act).
Although we believe these provisions benefit us by providing increased consistency in the application of Delaware law for the specified types of actions and proceedings, the provisions may have the effect of discouraging lawsuits against us or our directors and officers. Alternatively, if a court were to find the choice of forum provision contained in our amended and restated certificate of incorporation to be inapplicable or unenforceable in an action, we may incur additional costs associated with resolving such action in other jurisdictions, which could harm our business, financial condition, and operating results. For example, under the Securities Act, federal courts have concurrent jurisdiction over all suits brought to enforce any duty or liability created by the Securities Act, and investors cannot waive compliance with the federal securities laws and the rules and regulations thereunder. Any person or entity
130
purchasing or otherwise acquiring any interest in our shares of capital stock will be deemed to have notice of and consented to this exclusive forum provision, but will not be deemed to have waived our compliance with the federal securities laws and the rules and regulations thereunder.
Our ability to use our net operating loss carryforwards and certain other tax attributes may be limited.
Under the Tax Cuts and Jobs Act of 2017, as modified by the Coronavirus Aid, Relief, and Economic Stability Act (CARES Act), our federal net operating losses (NOLs) generated in tax years beginning after December 31, 2017, may be carried forward indefinitely, but the deductibility of such federal NOLs in tax years beginning after December 31, 2020, is limited to 80% of taxable income. It is uncertain if and to what extent various states will conform to the Tax Cuts and Jobs Act of 2017, or the CARES Act. In addition, under Sections 382 and 383 of the Internal Revenue Code of 1986, as amended, if a corporation undergoes an “ownership change,” generally defined as a greater than 50 percentage point change (by value) in its equity ownership by certain stockholders over a three-year period, the corporation’s ability to use its pre-change NOL and other pre-change tax attributes, such as research and development tax credits, to offset its post-change income or taxes may be limited. We may have experienced ownership changes in the past and may experience ownership changes as a result of subsequent shifts in our stock ownership, some of which are outside our control. As a result, our ability to use our pre-change NOLs and tax credits to offset post-change taxable income, if any, could be subject to limitations. Similar provisions of state tax law may also apply. In addition, at the state level, there may be periods during which the use of NOLs is suspended or otherwise limited, which could accelerate or permanently increase state taxes owed. For example, California recently imposed limits on the usability of California state NOLs and tax credits to offset California taxable income in tax years beginning after 2019 and before 2023. As a result, even if we attain profitability, we may be unable to use a material portion of our NOLs and tax credits.
General Risk Factors
Our quarterly operating results may fluctuate significantly or may fall below the expectations of investors or securities analysts, each of which may cause our stock price to fluctuate or decline.
We expect our operating results to be subject to quarterly fluctuations. Our net loss and other operating results will be affected by numerous factors, including:
|
|
• |
timing and variations in the level of expense related to the current or future development of our programs; |
|
|
• |
timing and status of enrollment for our future clinical trials; |
|
|
• |
changes or fluctuations in our stock price and market capitalization, which could impact the value of our contingent obligations and cause fluctuations in our operating expenses as a result of these non-cash adjustments; |
|
|
• |
impacts from the COVID-19 pandemic on us or third parties with which we collaborate or that we engage; |
|
|
• |
results of future clinical trials, or the addition or termination of such clinical trials or funding support by us or potential future partners; |
|
|
• |
our execution of any collaboration, licensing, or similar arrangements, and the timing of payments we may make or receive under such arrangements or the termination or modification of any such arrangements; |
|
|
• |
any intellectual property infringement, misappropriation, or violation lawsuit or opposition, interference, post-grant proceeding, or cancellation proceeding in which we may become involved; |
|
|
• |
additions and departures of key personnel; |
|
|
• |
strategic decisions by us or our competitors, such as acquisitions, divestitures, spin-offs, joint ventures, strategic investments, or changes in business strategy; |
|
|
• |
if any product candidate we may develop receives regulatory approval, the timing and terms of such approval and market acceptance and demand for such product candidate; |
|
|
• |
the timing and cost of establishing a sales, marketing, and distribution infrastructure to commercialize any products for which we may obtain marketing approval and intend to commercialize on our own or jointly with current or future collaborators; |
|
|
• |
regulatory developments affecting current or future product candidates or those of our competitors; |
|
|
• |
the amount of expense or gain associated with the change in value of the success payments and contingent consideration; and |
131
|
|
• |
changes in general market and economic conditions. |
If our quarterly operating results fall below the expectations of investors or securities analysts, the price of our common stock could decline substantially. Furthermore, any quarterly fluctuations in our operating results may, in turn, cause the price of our stock to fluctuate substantially. We believe that quarterly comparisons of our financial results are not necessarily meaningful and should not be relied upon as an indication of our future performance.
Our stock price may be volatile or may decline regardless of our operating performance, which may result in substantial losses for investors and may potentially subject us to securities class action litigation, which is expensive and could divert management’s attention.
The market price of our common stock may be highly volatile and may fluctuate substantially as a result of a variety of factors, some of which are related in complex ways. The market price of our common stock may fluctuate significantly in response to numerous factors, many of which are beyond our control, including the factors listed below and other factors describe in this “Risk Factors” section:
|
|
• |
the commencement of, enrollment in, or results of current and future preclinical studies and clinical trials we may conduct, or changes in the development status of our product candidates; |
|
|
• |
any delay in regulatory filings for our product candidates and any adverse development or perceived adverse development with respect to the applicable regulatory authority’s review of such filings, including the issuance by the FDA of a “refusal to file” letter or a request for additional information; |
|
|
• |
adverse results or delays in clinical trials; |
|
|
• |
our decision to initiate a preclinical study or clinical trial, not to initiate a preclinical study or clinical trial, or to terminate an existing preclinical study or clinical trial; |
|
|
• |
adverse actions taken by regulatory agencies with respect to our preclinical studies or clinical trials, manufacturing supply chain, or sales and marketing activities, including failure to receive regulatory approval of our product candidates; |
|
|
• |
changes in laws or regulations, including preclinical study or clinical trial requirements for regulatory approvals worldwide; |
|
|
• |
adverse changes to our relationship with manufacturers or suppliers; |
|
|
• |
manufacturing, supply, or distribution shortages; |
|
|
• |
our failure to successfully commercialize our product candidates; |
|
|
• |
changes in the structure of healthcare payment systems; |
|
|
• |
additions or departures of key scientific or management personnel; |
|
|
• |
unanticipated serious safety concerns related to the use of our product candidates; |
|
|
• |
disputes or other developments relating to proprietary rights, including patent rights, trade secrets, litigation matters, and our ability to obtain patent protection for our technologies or product candidates; |
|
|
• |
variations in our results of operations; |
|
|
• |
our cash position; |
|
|
• |
our failure to meet the estimates and projections of the investment community or that we may otherwise provide to the public; |
|
|
• |
publication of research reports about us or our industry, or ex vivo and in vivo cell engineering products in particular, or positive or negative recommendations or withdrawal of research coverage by securities analysts; |
|
|
• |
announcements made by us or our competitors about new product and service offerings, success or setbacks related to product or service offerings that exist or are under development, acquisitions, strategic relationships, joint ventures, or capital commitments; |
|
|
• |
our inability to establish collaborations, if needed; |
|
|
• |
our ability to effectively manage our growth; |
|
|
• |
the size and growth of our initial target markets; |
132
|
|
• |
changes in the market valuations of similar companies; |
|
|
• |
press reports, whether or not true, about our business; |
|
|
• |
sales or perceived potential sales of our common stock by us or our stockholders in the future; |
|
|
• |
overall fluctuations in the equity markets; |
|
|
• |
ineffectiveness of our internal controls; |
|
|
• |
changes in accounting practices or principles; |
|
|
• |
changes or developments in the global regulatory environment; |
|
|
• |
litigation involving us, our industry, or both, or investigations by regulators into our operations or those of our competitors; |
|
|
• |
general political and economic conditions, including geo-political and economic instability resulting from the recent escalation in conflict between Russia and Ukraine; and |
|
|
• |
other events or factors, many of which are beyond our control. |
In addition, the stock market in general, and biopharmaceutical companies in particular, have experienced extreme price and volume fluctuations that have often been unrelated or disproportionate to the operating performance of these companies. Broad market and industry factors may negatively affect the market price of our common stock, regardless of our actual operating performance. If the market price of our common stock does not exceed your purchase price, you may not realize any return on, and may lose some or all of, your investment.
In the past, securities class action litigation has often been instituted against companies following periods of volatility in the market price of a company’s securities. The market price of our common stock has fluctuated since our IPO and may continue in the future to be volatile. This type of litigation, if instituted, could result in substantial costs and a diversion of management’s attention and resources, which would harm our business, operating results, or financial condition.
We or the third parties upon whom we depend may be adversely affected by natural disasters, including earthquakes, fires, typhoons, and floods, public health epidemics, such as the ongoing COVID-19 pandemic, telecommunications or electrical failures, geo-political actions, including war and terrorism, political and economic instability, and other events beyond our control, and our business continuity and disaster recovery plans may not adequately protect us from a serious disaster.
We or our partners, CROs, CDMOs, or other service providers, may experience interruptions to our operations, including the conduct of our research and development programs, clinical trials, and manufacturing operations, due to natural disasters, including earthquakes, fires, typhoons, and floods, public health epidemics, such as the ongoing COVID-19 pandemic currently impacting countries worldwide, hardware, software, telecommunication or electrical failures, geo-political actions, including war and terrorism, or political and economic instability, which could significantly disrupt or harm our business.
Our corporate headquarters and other facilities, including the industrial space we lease on which we plan to build out and operate our manufacturing facility, are located in areas that have experienced significant natural disasters, including the San Francisco Bay Area and Seattle, Washington, each of which have experienced severe effects from wildfires and, in the case of the San Francisco Bay Area, severe earthquakes. We do not carry earthquake insurance. Earthquakes, wildfires, or other natural disasters could severely disrupt our operations, and could materially and adversely affect our business, financial condition, results of operations, and prospects. If a natural disaster, electrical failure, or other event occurs that prevents us from using all or a significant portion of our headquarters, damages critical infrastructure, or otherwise disrupts operations, it may be difficult or, in certain cases, impossible, for us to continue our business for a substantial period of time. For example, a prolonged electrical failure could result in damage to or destruction of materials that are critical for our research and manufacturing operations, including our master cell banks, which would delay the advancement of our programs and materially harm our business, operating results, prospects, or financial condition. In addition, a failure of our computing systems could result in the loss of research or preclinical data important to our research or development programs, interrupt the conduct of ongoing research, or otherwise impair our ability to operate, which could delay the advancement of our programs or cause us to incur costs to recover or reproduce lost data. In addition, if in the future a natural disaster, power outage, or other event occurred that prevented us from using all or a significant portion of our manufacturing facility, we may not be able to manufacture sufficient supply of our product candidates required to conduct our clinical trials or commercialize our products in accordance with our timelines or at all. The disaster recovery and business continuity plans we currently have in place are limited and are unlikely to prove adequate in the event of a serious disaster or similar event. We may incur substantial expenses as a result of the limited nature of our disaster recovery and business continuity plans, which, together with our lack of earthquake insurance in particular, could have a material adverse effect on our business.
133
Integral parties in our supply chain are similarly vulnerable to natural disasters or other sudden, unforeseen, and severe adverse events. In addition, our supply chain is vulnerable to changes in the geo-political and economic climate, including changes in relationships between the United States and countries from which we may need to source materials and other resources necessary for the preclinical evaluation of our product candidates, including animal models, and specifically non-human primate models, or to manufacture our product candidates, including raw and intermediate materials and consumables. If any such event or change were to affect our supply chain, it could have a material adverse effect on our business.
Furthermore, geo-political actions, and the resulting political and economic instability, could negatively impact our operations. For example, in late February 2022, Russia initiated significant military action against Ukraine. In response, the United States and certain other countries imposed significant sanctions and trade actions against Russia and could impose further sanctions, trade restrictions, and other retaliatory actions if the conflict continues or worsens. It is not possible to predict the broader consequences of the conflict, including related geo-political tensions, and the measures and retaliatory actions that will be taken by the United States and other countries in respect thereof, as well as any countermeasures or retaliatory actions Russia may take in response, are likely to cause regional instability and geo-political shifts and could materially adversely affect global trade, currency exchange rates, regional economies, and the global economy. While it is difficult to anticipate the impact of any of the foregoing on our company in particular, the conflict and actions taken in response to the conflict could increase our costs, disrupt our supply chain, impair our ability to raise or access additional capital when needed on acceptable terms, if at all, or otherwise adversely affect our business, financial condition, and results of operations.
We are subject to United States and certain foreign export and import controls, sanctions, embargoes, anti-corruption laws, and anti-money laundering laws and regulations. Compliance with these legal standards could impair our ability to compete in domestic and international markets. We could face criminal liability and other serious consequences for violations, which would harm our business.
We are subject to export control and import laws and regulations, including the United States Export Administration Regulations, United States Customs regulations, various economic and trade sanctions regulations administered by the United States Treasury Department’s Office of Foreign Assets Controls, the United States Foreign Corrupt Practices Act of 1977, as amended (FCPA), the United States domestic bribery statute contained in 18 U.S.C. § 201, the United States Travel Act, the USA PATRIOT Act, and other state and national anti-bribery and anti-money laundering laws in the countries in which we conduct activities. Anti-corruption laws are interpreted broadly and prohibit companies and their employees, agents, contractors, and other collaborators from authorizing, promising, offering, or providing, directly or indirectly, improper payments or anything else of value to recipients in the public or private sector. We may engage third parties to sell any products for which we receive regulatory approval outside the United States, to conduct clinical trials, and/or to obtain necessary permits, licenses, patent registrations, and other regulatory approvals. We have direct or indirect interactions with officials and employees of government agencies or government-affiliated hospitals, universities, and other organizations. We can be held liable for the corrupt or other illegal activities of our employees, agents, contractors, and other collaborators, even if we do not explicitly authorize or have actual knowledge of such activities. Any violations of the laws and regulations described above may result in substantial civil and criminal fines and penalties, imprisonment, the loss of export or import privileges, debarment, tax reassessments, breach of contract and fraud litigation, reputational harm, and other consequences.
The withdrawal of the United Kingdom from the European Union, commonly referred to as “Brexit,” may adversely impact our ability to obtain regulatory approvals of our product candidates in the United Kingdom or European Union, result in restrictions or imposition of taxes and duties for importing our product candidates into the United Kingdom or European Union, and may require us to incur additional expenses in order to develop, manufacture, and commercialize our product candidates in the United Kingdom or European Union.
Following the result of a referendum in 2016, the United Kingdom (UK) left the European Union (EU) on January 31, 2020, commonly referred to as “Brexit.” Pursuant to the formal withdrawal arrangements agreed between the United Kingdom and the EU, the United Kingdom was subject to a transition period until December 31, 2020 (the Transition Period), during which time EU rules continued to apply. Negotiations between the United Kingdom and the EU continue in relation to the customs and trading relationship between the United Kingdom and the EU following the expiry of the Transition Period.
Since a significant proportion of the regulatory framework in the United Kingdom applicable to our business and our product candidates is derived from EU directives and regulations, Brexit could materially impact the regulatory regime with respect to the development, manufacture, importation, approval, and commercialization of our product candidates in the United Kingdom or the EU. For example, as a result of the uncertainty surrounding Brexit, the EMA relocated to Amsterdam from London. Following the Transition Period, the United Kingdom is no longer covered by the centralized procedures for obtaining EU-wide marketing authorization from the EMA and, unless a specific agreement is entered into, a separate process for authorization of drug products, including our product candidates, will be required in the United Kingdom, the potential process for which is currently unclear. Any delay in obtaining, or an inability to obtain, any marketing approvals, as a result of Brexit or otherwise, would prevent us from
134
commercializing our product candidates in the United Kingdom or the EU and restrict our ability to generate revenue and achieve and sustain profitability. In addition, we may be required to pay taxes or duties or be subjected to other hurdles in connection with the importation of our product candidates into the EU, or we may incur expenses in establishing a manufacturing facility in the EU in order to circumvent such hurdles. If any of these outcomes occur, we may be forced to restrict or delay efforts to seek regulatory approval in the United Kingdom or the EU for our product candidates or incur significant additional expenses to operate our business, which could significantly and materially harm or delay our ability to generate revenues or achieve profitability. Any further changes in international trade, tariff, and import/export regulations as a result of Brexit or otherwise may impose unexpected duty costs or other non-tariff barriers on us. These developments, or the perception that any of them could occur, may significantly reduce global trade and, in particular, trade between the impacted nations and the United Kingdom.
Since the beginning of 2021,when the Transition Period expired, we have been required to comply with the GDPR as well as the UK GDPR. Each regime has the ability to impose fines of up to the greater of €20 million (£17.5 million) or 4% of global turnover for non-compliance. The relationship between the UK and the EU in relation to transfers of personal data from the EU to the UK is not fully settled by the Brexit Trade and Cooperation Agreement (TCA). Instead, the TCA establishes a four- to six-month grace period during which transfers of personal data from the EU to the UK can continue without additional safeguards, provided that the UK maintains its pre-TCA data protection laws. During this time, the European Commission may adopt a UK adequacy decision which may be relied upon by organizations for EU to UK personal data transfers, but if no UK adequacy decision is adopted, the UK will be considered a third country at the end of the grace period and we will be required to implement additional safeguards for personal data transfers—some of which are subject currently being scrutinized or challenged—which could lead to additional costs and increase our overall risk exposure.
If securities or industry analysts either do not publish research about us or publish inaccurate or unfavorable research about us, our business, our market, or our competitors, or if they change their recommendations regarding our common stock adversely, the trading price or trading volume of our common stock could decline.
The trading market for our common stock will be influenced in part by the research and reports that securities or industry analysts may publish about us, our business, our market, or our competitors. If one or more of these analysts initiate research with an unfavorable rating or downgrade our common stock, provide a more favorable recommendation about our competitors, or publish inaccurate or unfavorable research about our business, our common stock price would likely decline. If any analyst who may cover us were to cease such coverage or fail to regularly publish reports on us, we could lose visibility in the financial markets, which could cause the trading price or trading volume of our common stock to decline.
We are an emerging growth company, and any decision on our part to comply only with certain reduced reporting and disclosure requirements applicable to emerging growth companies could make our common stock less attractive to investors.
We are an “emerging growth company” as defined in the JOBS Act, and, for as long as we continue to be an emerging growth company, we may choose to take advantage of exemptions from various reporting requirements applicable to other public companies but not to emerging growth companies, including:
|
|
• |
not being required to have our independent registered public accounting firm audit our internal control over financial reporting under Section 404 of the Sarbanes-Oxley Act of 2002 (the Sarbanes-Oxley Act); |
|
|
• |
reduced disclosure obligations regarding executive compensation in our periodic reports and annual report on Form 10-K; and |
|
|
• |
exemptions from the requirements of holding non-binding advisory votes on executive compensation and stockholder approval of any golden parachute payments not previously approved. |
Our status as an emerging growth company will end as soon as any of the following takes place:
|
|
• |
the last day of the fiscal year in which we have more than $1.07 billion in annual revenue; |
|
|
• |
the date we qualify as a “large accelerated filer,” with at least $700 million of equity securities held by non-affiliates; |
|
|
• |
the date on which we have issued, in any three-year period, more than $1.0 billion in non-convertible debt securities; or |
the last day of the fiscal year ending after the fifth anniversary of the completion of our initial public offering, which is December 31, 2026.
We cannot predict if investors will find our common stock less attractive as a result of our decision to rely on any of the exemptions afforded to emerging growth companies. If some investors find our common stock less attractive because we rely on any of these exemptions, there may be a less active trading market for our common stock and the market price of our common stock may be more volatile.
135
Under the JOBS Act, emerging growth companies can also delay adopting new or revised accounting standards until such time as those standards apply to private companies. We have elected to use this extended transition period for any new or revised accounting standards during the period in which we remain an emerging growth company (or we affirmatively and irrevocably opted out of the extended transition period); however, we may adopt certain new or revised accounting standards early. As a result, these financial statements may not be comparable to companies that comply with the new or revised accounting pronouncements as of public company effective dates.
The requirements of being a public company may strain our resources, result in an increased risk of litigation, and divert management’s attention.
As a public company, we are subject to the reporting requirements of the Exchange Act, the Sarbanes-Oxley Act, the Dodd-Frank Wall Street Reform and Consumer Protection Act (the Dodd-Frank Act), the listing requirements of Nasdaq, and other applicable securities rules and regulations. Complying with these rules and regulations has increased and will increase our legal and financial compliance costs, make some activities more difficult, time-consuming, or costly, and increase demand on our systems and resources. The Exchange Act requires, among other things, that we file annual, quarterly, and current reports with respect to our business and operating results. The Sarbanes-Oxley Act requires, among other things, that we maintain effective disclosure controls and procedures and internal control over financial reporting. We are required to disclose changes made in our internal control and procedures on a quarterly basis. In order to maintain and, if required, improve our disclosure controls and procedures and internal control over financial reporting to meet the requirements of the Sarbanes-Oxley Act, significant resources and management oversight may be required. As a result, management’s attention may be diverted from other business concerns, which could adversely affect our business and operating results. We may also need to hire additional employees or engage outside consultants to comply with these requirements, which will increase our costs and expenses.
In addition, changing laws, regulations, and standards relating to corporate governance and public disclosure are creating uncertainty for public companies, increasing legal and financial compliance costs, and making some activities more time-consuming. These laws, regulations, and standards are subject to varying interpretations, in many cases due to their lack of specificity and, as a result, their application in practice may evolve over time as new guidance is provided by regulatory and governing bodies. This could result in continuing uncertainty regarding compliance matters and higher costs necessitated by ongoing revisions to disclosure and governance practices. We intend to invest resources to comply with evolving laws, regulations, and standards, and this investment may result in increased general and administrative expenses and a diversion of management’s time and attention from potential revenue-generating activities to compliance activities. If our efforts to comply with new laws, regulations, and standards fail to meet the requirements of the applicable regulatory or governing bodies due to ambiguities related to their application in practice, regulatory authorities may initiate legal proceedings against us, and our business may be adversely affected.
These new rules and regulations may make it more expensive for us to obtain director and officer liability insurance and, in the future, we may be required to accept reduced coverage or incur substantially higher costs to obtain coverage. These factors could also make it more difficult for us to attract and retain qualified members of our Board, particularly to serve on our audit committee and compensation committee, and qualified executive officers. Additionally, the dramatic increase in the cost of such insurance may cause us to opt for lower overall policy limits or to forgo insurance that we may otherwise rely on to cover defense costs, settlements, and damages awarded to plaintiffs in connection with any securities litigation.
By disclosing information in the periodic filings required of a public company, our business and financial condition are more visible, which we believe may result in threatened or actual litigation, including by competitors and other third parties. If those claims are successful, our business could be seriously harmed. Even if the claims do not result in litigation or are resolved in our favor, the time and resources needed to resolve them could divert our management’s resources and seriously harm our business.
If we fail to maintain proper and effective internal controls over financial reporting, our ability to produce accurate and timely financial statements could be impaired.
Pursuant to Section 404 of the Sarbanes-Oxley Act, our management is required to report upon the effectiveness of our internal control over financial reporting. When we lose our status as an “emerging growth company” and become an “accelerated filer” or a “large accelerated filer,” our independent registered public accounting firm will be required to attest to the effectiveness of our internal control over financial reporting. The rules governing the standards that must be met for management to assess our internal control over financial reporting are complex and require significant documentation, testing, and possible remediation. To comply with the requirements of being a reporting company under the Exchange Act, we will need to implement additional financial and management controls, reporting systems, procedures, and hire additional accounting and finance staff.
We cannot guarantee that there will not be material weaknesses or significant deficiencies in our internal control over financial reporting in the future. Any failure to maintain internal control over financial reporting could severely inhibit our ability to accurately report our financial condition, results of operations, or cash flows. If we are unable to conclude that our internal control over financial
136
reporting is effective, or if our independent registered public accounting firm determines we have a material weakness or significant deficiency in our internal control over financial reporting, investors may lose confidence in the accuracy and completeness of our financial reports, the market price of our common stock could decline, and we could be subject to sanctions or investigations by Nasdaq, the SEC, or other regulatory authorities. Failure to remedy any material weakness in our internal control over financial reporting, or to implement or maintain other effective control systems required of public companies, could also restrict our future access to the capital markets.
Our disclosure controls and procedures may not prevent or detect all errors or acts of fraud.
We are subject to the periodic reporting requirements of the Exchange Act. We must design our disclosure controls and procedures to reasonably assure that information we must disclose in reports we file or submit under the Exchange Act is accumulated and communicated to management, and recorded, processed, summarized, and reported within the time periods specified in the rules and forms of the SEC. We believe that any disclosure controls and procedures or internal controls and procedures, no matter how well-conceived and operated, can provide only reasonable, not absolute, assurance that the objectives of the control system are met. These inherent limitations include the realities that judgments in decision-making can be faulty, and that breakdowns can occur because of simple error or mistake. For example, our directors or executive officers could inadvertently fail to disclose a new relationship or arrangement with a related party, which could cause us to fail to make a required related party transaction disclosure. Additionally, controls can be circumvented by the individual acts of some persons, by collusion of two or more people, or by an unauthorized override of the controls. Accordingly, because of the inherent limitations in our control system, misstatements due to error or fraud may occur and not be detected.
137
Item 1B. Unresolved Staff Comments.
None.
Item 2. Properties.
Washington
Our corporate headquarters are located in Seattle, Washington, where we lease 25,898 square feet of office and laboratory space pursuant to a lease agreement that commenced in March 2019 and expires in December 2026, and includes an option to extend for an additional five-year term. We sublease an additional 22,188 square feet of office and laboratory space in the same building, pursuant to a sublease agreement which commenced in September 2020 and expires in April 2028.
California
We occupy 66,075 square feet of office and laboratory space in South San Francisco, California, pursuant to a lease agreement that commenced in December 2019 expires in April 2030, and includes the option to extend for an additional five-year term. In July 2021, we entered into a lease agreement for 163,193 square feet of industrial space located in Fremont, California, which will be used for the construction of a GMP manufacturing facility. The lease agreement has an initial term of ten years expiring in November 2031 and includes an option to extend for up to two additional five-year terms.
Massachusetts
We lease 24,386 square feet of office and laboratory space in Cambridge, Massachusetts pursuant to a lease agreement that commenced in March 2019 and expires in June 2027 and sublease an additional 31,563 square feet of similar space in an adjacent building pursuant to a sublease agreement that commenced in January 2020 and expires in February 2028.
We believe that our existing facilities are sufficient for our near-term needs but expect to need additional space as we grow. We believe that suitable additional alternative spaces will be available in the future on commercially reasonable terms, if required.
Item 3. Legal Proceedings.
We are not currently a party to any material legal proceedings. From time to time, we may, however, in the ordinary course of business face various claims brought by third parties, including claims relating to employment matters and the safety and efficacy of our products, and we may, from time to time, make claims or take legal action to assert our rights, including action relating to our intellectual property rights. Any of these claims could subject us to costly litigation. While we generally believe that we have adequate insurance to cover many different types of liabilities from third party claims, our insurance carriers may deny coverage, may be inadequately capitalized to pay on valid claims, or our policy limits may be inadequate to fully satisfy any damage awards or settlements. If this were to happen, the payment of any such awards could have a material adverse effect on our operations, cash flows and financial position. Additionally, any such claims, whether or not successful, could damage our reputation and business.
Item 4. Mine Safety Disclosures.
Not applicable.
138
PART II
Item 5. Market for Registrant’s Common Equity, Related Stockholder Matters and Issuer Purchases of Equity Securities.
Market Information
Our common stock has been listed on The Nasdaq Global Select Market under the symbol “SANA” since February 4, 2021. Prior to that date, there was no public trading market for our common stock.
Holders
As of March 11, 2022, there were approximately 108 holders of record of our common stock. The actual number of stockholders is greater than this number of record holders, and includes stockholders who are beneficial owners, but whose shares are held in street name by brokers and other nominees. This number of holders of record also does not include stockholders whose shares may be held in trust by other entities.
Dividend Policy
We have not declared or paid cash dividends on our capital stock since our inception. We intend to retain future earnings, if any, to finance the operation and expansion of our business and do not anticipate paying any cash dividends to holders of common stock in the foreseeable future.
Stock Performance Graph
The following performance graph and related information shall not be deemed to be “soliciting material” or to be “filed” with the SEC, for purposes of Section 18 of the Securities Exchange Act of 1934, as amended, or the Exchange Act, nor shall such information be incorporated by reference into any future filing under the Exchange Act or Securities Act of 1933, as amended, or the Securities Act, except to the extent that we specifically incorporate it by reference into such filing.
The graph set forth below compares the cumulative total stockholder return on our shares of common stock between February 5, 2021 (the date of our initial public offering) and December 31, 2021, with the cumulative total return of (a) the Nasdaq Biotechnology Index and (b) the Nasdaq Composite Index, over the same period. This graph assumes the investment of $100 on February 5, 2021 in our common stock, the Nasdaq Biotechnology Index and the Nasdaq Composite Index and assumes the reinvestment of dividends, if any. The graph uses the closing sales price of our common stock of $39.12 per share on February 5, 2021 as the initial value of our common stock and not the initial offering price to the public of $25.00 per share. The comparisons shown in the graph below are based upon historical data. The stock price performance included in this graph is not necessarily indicative of future stock price performance.
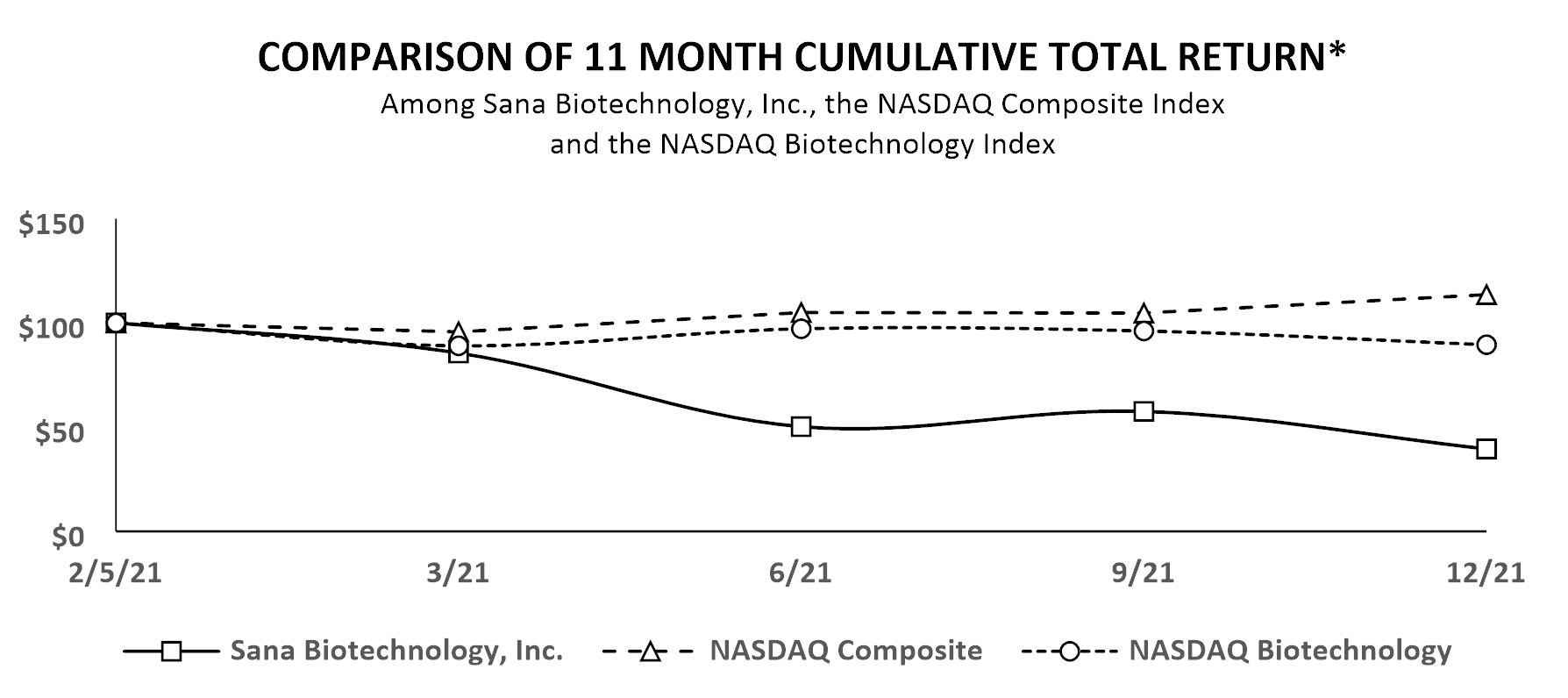
*$100 invested on 2/5/21 in stock or index, including reinvestment of dividends.
Fiscal year ending December 31.
139
Securities Authorized for Issuance Under Equity Compensation Plans
Information about our equity compensation plans is incorporated herein by reference to Item 12 of Part III of this Annual Report.
Recent Sales of Unregistered Securities
None.
Use of Proceeds from our Initial Public Offering of Common Stock
On February 8, 2021, we closed an initial public offering (IPO) and issued and sold 27,025,000 shares of our common stock, including 3,525,000 shares of common stock sold pursuant to the underwriters’ full exercise of their option to purchase additional shares, at a public offering price of $25.00 per share, for aggregate gross proceeds of $675.6 million. All of the shares issued and sold in the IPO were registered under the Securities Act pursuant to a Registration Statement on Form S-1 (File No. 333-252061), which was declared effective by the SEC on February 3, 2021, and a Registration Statement on Form S-1 MEF (File No. 333-252706) filed pursuant to Rule 462(b) of the Securities Act. Morgan Stanley & Co. LLC, Goldman Sachs & Co. LLS, J.P. Morgan Securities LLC, and BofA Securities, Inc. acted as joint bookrunning managers of the IPO and as representatives of the underwriters. No offering expenses were paid directly or indirectly to any of our directors or officers (or their associates) or persons owning 10.0% or more of any class of our equity securities or to any other affiliates. We are holding a significant portion of the balance of the net proceeds from the offering in money market funds and short-term investments. There has been no material change in the planned use of proceeds from the IPO from that described in the prospectus filed with the SEC pursuant to Rule 424(b)(4) under the Securities Act, which was declared effective on February 3, 2021. We invested the funds received in cash equivalents and other marketable securities in accordance with our investment policy.
Issuer Purchases of Equity Securities
None.
Item 6. Reserved
140
Item 7. Management’s Discussion and Analysis of Financial Condition and Results of Operations.
You should read the following discussion and analysis of our financial condition and results of operations together with the section titled “Selected Financial Data,” and our audited consolidated financial statements and the related notes included elsewhere in this Annual Report. This discussion and analysis and other parts of this Annual Report contain forward-looking statements that are based upon current beliefs, plans, and expectations related to future events and our future financial performance that involve risks, uncertainties, and assumptions, such as statements regarding our intentions, plans, objectives, and expectations for our business. Our actual results and the timing of selected events could differ materially from those described in or implied by these forward-looking statements as a result of numerous factors, including those set forth in the section of this Annual Report titled “Risk Factors.” See also the section of this Annual Report titled “Special Note Regarding Forward-Looking Statements.”
Overview
We were founded on the belief that engineered cells will be one of the most important transformations in medicine over the next several decades. The burden of diseases that can be addressed at their root cause through engineered cells is significant. We view engineered cells as having the potential to be as therapeutically disruptive as biologics to clinical practice. Our long-term aspirations are to be able to control or modify any gene in the body, to replace any cell that is damaged or missing, and to markedly improve access to cellular and gene-based medicines. We have brought together an experienced group of scientists, engineers, and company builders and combined them with the necessary technologies to move this vision forward. We are developing ex vivo and in vivo cell engineering platforms to revolutionize treatment across a broad array of therapeutic areas with unmet treatment needs, including oncology, diabetes, central nervous system disorders, cardiovascular diseases, and genetic disorders, among others. Our platform progress, broad capabilities, and strong balance sheet enable us to execute on a broad vision, with a goal of submitting our first INDs in 2022, with the opportunity to file multiple INDs per year beyond 2022.
Frequently in disease, cells are damaged or missing entirely, and an effective therapy needs to replace the entire cell, an approach referred to as cell therapy or ex vivo cell engineering. A successful therapeutic requires an ability to manufacture cells at scale that engraft, function, and have the necessary persistence in the body. Of these, long-term persistence related to overcoming immunologic rejection of another person’s cells has been the most challenging, which has led many to focus on autologous, or a patient’s own, cells as the therapeutic source. However, autologous therapies require a complex process of harvesting cells from the patients, manipulating them outside the body, and returning them to the patient. Products utilizing this approach have had to manage significant challenges such as scalability, product variability, product quality, cost, patient accessibility, and a limited number of cell types being amenable to this approach. Given these limitations, rather than utilizing autologous cells to overcome immune rejection, we have invested in creating hypoimmune cells that can “hide” from the patient’s immune system. We are striving to make therapies utilizing pluripotent stem cells with our hypoimmune genetic modifications as the starting material, which we then differentiate into a specific cell type, such as a pancreatic beta cell, before treating the patient. Additionally, for cell types for which effective differentiation protocols from a stem cell have not yet been developed, such as T cells, instead of starting from a pluripotent stem cell, we can utilize an allogeneic cell, differentiated cells sourced from a donor, as the starting material to which we then apply our hypoimmune genetic modifications.
The process of repairing and controlling genes in the body, referred to as gene therapy or in vivo cell engineering, requires in vivo delivery of a therapeutic payload and modification of the genome. There are multiple methods available to modify the genome, but limited ability to deliver therapeutic payloads in vivo. Thus, delivery of a therapeutic payload is at the core of our strategic focus, with our ultimate goal being the delivery of any payload to any cell in a specific and repeatable way. Our initial effort is on cell-specific delivery and increasing the diversity and size of payloads. Using our fusogen technology, we have shown in preclinical studies that we can specifically target numerous cell surface receptors that, when combined with delivery vehicles to form fusosomes, allow cell-specific delivery across multiple different cell types. We have initially chosen to focus this technology on delivering payloads to T cells, hepatocytes, and hematopoietic stem cells.
We believe the time is right to develop engineered cell therapies across a broad range of therapeutic areas. Substantial progress in the understanding of genetics, gene editing, gene control, protein engineering, stem cell biology, immunology, process analytics, and computational biology have converged to create an opportunity to markedly increase the breadth and depth of the potential impact of genetic and cellular medicines. We are focused on creating transformative ex vivo and in vivo engineered cell therapies across a
141
range of therapeutic areas. We are in the early stages of development across a broad pipeline of product candidates, all of which are currently in the preclinical stage of development and are summarized below:
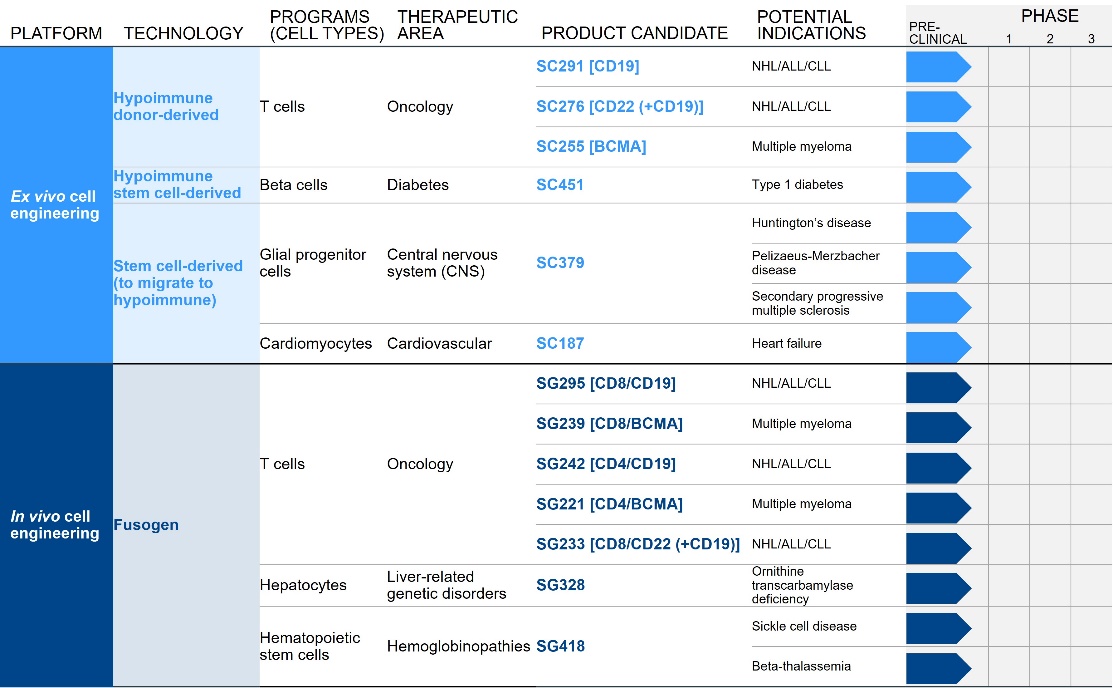
We continue to make progress on developing our cell engineering platforms and advancing our product candidates through preclinical development and towards potential IND submissions. Given the depth and breadth of our portfolio, we expect to assess and prioritize our programs on an ongoing basis based on various factors, including internal and external opportunities and constraints, which may result in our decision to advance certain programs ahead or instead of others. As certain of our product candidates advance towards potential IND submissions, we are conducting GLP toxicity studies and establishing necessary scale-up for our manufacturing processes. Our goal is to file INDs in 2022 for our hypoimmune allogeneic CD19 CAR T (SC291) and our in vivo CD19 CAR T (SG295) product candidates. For details regarding our product candidates, see the section titled “Business— Overview” in Part I, Item 1 included elsewhere in this Annual Report.
Our ex vivo and in vivo technologies represent an aggregation of years of innovation and technology from multiple academic institutions and companies, including our ex vivo cell engineering programs focused on replacing damaged cells in the heart and certain brain disorders acquired from Cytocardia Inc. (Cytocardia) and Oscine Corp. (Oscine), respectively, hypoimmune technology licensed from the President and Fellows of Harvard College (Harvard) and The Regents of the University of California (UCSF), fusogen technology acquired from Cobalt Biomedicines Inc. (Cobalt), and genome editing technology licensed from Beam Therapeutics Inc. (Beam), among others. For details regarding these acquisitions and license and collaboration agreements, see Note 3, Acquisitions and Note 4, License and collaboration agreements, to our consolidated financial statements included in this Annual Report, as well as the section titled “Business— Key Intellectual Property Agreements” in Part I, Item 1 included elsewhere in this Annual Report.
We were incorporated in July 2018, and our operations to date have included developing our ex vivo and in vivo cell engineering platforms, identifying and developing potential product candidates, executing preclinical studies, establishing manufacturing capabilities, acquiring technology, organizing and staffing the company, business planning, establishing our intellectual property portfolio, raising capital, and providing general and administrative support for these operations. All of our programs are currently in the development stage, and we do not have any products approved for sale. Since our inception, we have incurred net losses each year. Our net losses for the years ended December 31, 2021, 2020, and 2019 were $355.9 million, $285.3 million, and $130.8 million, respectively. As of December 31, 2021, we had an accumulated deficit of $785.4 million. Our net losses resulted primarily from our research and development programs, and, to a lesser extent, general and administrative costs associated with our operations. In addition, as of December 31, 2021, the accumulated deficit of $785.4 million includes non-cash charges of $100.1 million and $102.5 million related to the revaluation of the success payment liabilities and contingent consideration, respectively.
142
In February 2021, we completed our initial public offering (IPO) and issued 27.0 million shares of our common stock, including 3.5 million shares pursuant to the full exercise of the underwriters’ option to purchase additional shares, at a price of $25.00 per share and received net proceeds of $626.4 million. Prior to the IPO, we funded our operations from the issuance and sale of our convertible preferred stock, raising an aggregate of $705.5 million in gross proceeds. As of December 31, 2021, we had cash, cash equivalents, and marketable securities of $746.9 million. Based on our current operating plan, we believe that our existing cash, cash equivalents, and marketable securities will be sufficient to meet our working capital and capital expenditure needs for at least the next 12 months. We anticipate that our expenses and operating losses will increase substantially for the foreseeable future. The expected increase in expenses will be driven in large part by our ongoing activities if and as we: continue to advance our ex vivo and in vivo cell engineering platforms; continue preclinical development of our current and future product candidates and initiate additional preclinical studies; commence clinical studies of our current and future product candidates; establish our manufacturing capabilities, including developing our contract development and manufacturing relationships and building our internal manufacturing facility; acquire and license technologies aligned with our ex vivo and in vivo cell engineering platforms; seek regulatory approval of our current and future product candidates; expand our operational, financial, and management systems; increase personnel, including personnel to support our preclinical and clinical development, manufacturing, and commercialization efforts; continue to develop, grow, prosecute, and defend our intellectual property portfolio; and incur additional legal, accounting, or other expenses in operating our business, including the costs associated with operating as a public company.
We are investing early in building world class capabilities in key areas of manufacturing sciences and operations, including development of our ex vivo and in vivo cell engineering platforms, product characterization, and process analytics from the time candidates are in early research phases. Our investments also include scaled research solutions, scaled infrastructure, and novel technologies to improve efficiency, characterization, and scalability of manufacturing, including establishing our internal manufacturing facility.
We anticipate that we will need to raise additional financing in the future to fund our operations, including the commercialization of any approved product candidates. Until we can generate significant product revenue, if ever, we expect to finance our operations with our existing cash, cash equivalents, and marketable securities, the proceeds of any future equity or debt financings, and upfront, milestone, and royalty payments, if any, received under future license or collaboration agreements. We may not be able to raise additional capital on terms that are acceptable to us or at all. If we are unable to raise additional capital when desired, our business, results of operations, and financial condition would be adversely affected.
COVID-19 business update
The global COVID-19 pandemic continues to evolve rapidly, and we continue to monitor it closely. The extent of the impact of the ongoing COVID-19 pandemic on our business, operations, and clinical development timelines and plans remains uncertain and will depend on certain developments, including the duration of the COVID-19 pandemic and spread of COVID-19, and the pandemic’s impact on our ability to build out and operationalize our internal manufacturing facility, expand our laboratory space, and enroll patients in clinical trials, and the impact of the pandemic on our clinical trial sites, contract research organizations (CROs), contract manufacturing organizations, suppliers of key materials and supplies, including raw materials, consumables, and other equipment necessary to manufacture our product candidates, and other third parties with whom we do business, as well as its impact on regulatory authorities and our key scientific and management personnel. We have experienced modest delays in our discovery and development activities as a result of the COVID-19 pandemic, primarily due to temporary and partial shutdowns at certain of our CROs and academic institutions that have since resumed operations, and stay-at-home orders in Washington, California, and Massachusetts, where our operations are located. However, to the extent possible, we are conducting business as usual, with necessary or advisable modifications to employee travel and most of our non-laboratory employees primarily working remotely. We continue to actively monitor the situation related to COVID-19 and may take further actions that alter our operations, including those that may be required by federal, state, or local authorities, or that we determine are in the best interests of our employees and other third parties with which we do business.
Acquisitions
We have completed various acquisitions since inception. For details regarding our acquisitions, see the section titled “Business—Key Intellectual Property Agreements” and Note 3, Acquisitions, to our consolidated financial statements included elsewhere in this Annual Report.
License and collaboration agreements
We have entered into license and collaboration agreements with various third parties. For details regarding these agreements, see the section titled “Business— Key Intellectual Property Agreements” and Note 4, License and collaboration agreements, to our consolidated financial statements included elsewhere in this Annual Report.
143
Success payments and contingent consideration
Cobalt success payment and contingent consideration
Pursuant to the terms of the Cobalt acquisition agreement, we have an obligation to pay to certain former Cobalt stockholders’ contingent consideration (Cobalt Contingent Consideration) of up to an aggregate of $500.0 million upon our achievement of certain pre-specified development milestones and a success payment (Cobalt Success Payment) of up to $500.0 million payable in cash or stock. The Cobalt Success Payment is payable if, at pre-determined valuation measurement dates, including the closing of our IPO and periodically thereafter, our market capitalization equals or exceeds $8.1 billion and we are advancing a program based on the fusogen technology in a clinical trial pursuant to an IND, or have filed for, or received approval for, a biologics license application (BLA) or new drug application (NDA). As of December 31, 2021, a Cobalt Success Payment had not been triggered. In addition to our IPO, a valuation measurement date would be triggered upon a change of control if at least one of our programs based on the fusogen technology is the subject of an active research program at the time of such change of control. If there is a change of control and our market capitalization is below $8.1 billion as of the date of such change of control, the amount of the potential Cobalt Success Payment will decrease, and the amount of potential Cobalt Contingent Consideration will increase. See Note 3, Acquisitions to our consolidated financial statements included elsewhere in this Annual Report for details on the amount of the potential Cobalt Success Payment and potential Cobalt Contingent Consideration if there is a change of control based on various thresholds for our market capitalization on such change of control date.
As of December 31, 2021 and 2020, the estimated fair value of the Cobalt Success Payment liability was $88.3 million and $64.7 million, respectively, and were recorded in long-term liabilities in the consolidated balance sheets. In connection with the change in the estimated fair value of the Cobalt Success Payment, we recognized expenses of $23.6 million, $62.3 million, and an immaterial amount, respectively, for the years ended December 31, 2021, 2020, and 2019.
As of December 31, 2021, the estimated fair value of the Cobalt Contingent Consideration was $153.7 million, of which $51.4 million was recorded in short-term liabilities and $102.3 million was recorded in long-term liabilities in the consolidated balance sheet. As of December 31, 2020, the estimated fair value of the Cobalt Contingent Consideration of $121.9 million was recorded in long-term liabilities in the consolidated balance sheet. In connection with the change in the estimated fair value of the Cobalt Contingent Consideration, we recognized expenses of $31.8 million, $52.8 million, and $17.9 million, respectively, for the years ended December 31, 2021, 2020, and 2019. See the subsections below titled “—Success payments” and “—Contingent consideration” for more information on the accounting treatment of the Cobalt Success Payment and Cobalt Contingent Consideration.
Harvard success payments
Pursuant to the terms of the Harvard agreement, we may be required to make up to an aggregate of $175.0 million in success payments to Harvard (Harvard Success Payments), payable in cash, based on increases in the per share fair market value of our common stock. The potential Harvard Success Payments are based on multiples of increased value ranging from 5x to 40x based on a comparison of the per share fair market value of our common stock relative to the original issuance price of $4.00 per share at pre-determined valuation measurement dates. The Harvard Success Payments can be achieved over a maximum of 12 years from the effective date of the agreement. See Note 4, License and collaboration agreements to our consolidated financial statements included elsewhere in this Annual Report for more details on the various per share common stock values that trigger a Harvard Success Payment.
The first valuation measurement date occurred in February 2022 on the one-year anniversary of our IPO. The threshold was not met, and therefore no payment was made as of the measurement date. Additional valuation measurement dates are triggered by certain events, including a merger, an asset sale, the sale of the majority of the shares held by Series A convertible preferred stockholders, and the last day of the term of the Harvard Success Payments. If a higher success payment tier is met at the same time a lower tier is met, both tiers will be owed. Any previous Harvard Success Payments made are credited against the Harvard Success Payment owed as of any valuation measurement date so that Harvard does not receive multiple success payments in connection with the same threshold.
As of December 31, 2021 and 2020, the estimated fair value of the Harvard Success Payment liability was $14.2 million and $11.8 million, respectively. As of December 31, 2021 and 2020, $5.0 million and $0, respectively, were recorded in short-term liabilities, and $9.2 million and $11.8 million, respectively, were recorded in long-term liabilities in the consolidated balance sheet. In connection with the change in the estimated fair value of the Harvard Success Payment liability, we recognized expenses of $2.4 million, $9.9 million, and $1.9 million, respectively, for the years ended December 31, 2021, 2020, and 2019. See the subsection below titled “—Success payments” for more information on the accounting treatment of the Harvard Success Payments.
144
Components of operating results
Operating expenses
Research and development
To date, research and development expenses have related primarily to discovery and development of our platform technology and product candidates. Research and development expenses are recognized as incurred, and payments made prior to the receipt of goods or services to be used in research and development are recorded as prepaid expenses, until the goods or services are received.
Research and development expenses consist of personnel-related costs, including salaries, benefits, and non-cash stock-based compensation, external research and development expenses incurred under arrangements with third parties, costs for laboratory supplies, costs to acquire and license technologies aligned with our goal of translating engineered cells to medicines, facility and other allocated expenses, including rent, depreciation, and allocated overhead costs, and other research and development expenses. The timing and amount of costs to acquire and license technologies in the future cannot be reliably estimated and may fluctuate from quarter to quarter and year to year.
We deploy our employee and infrastructure resources across multiple research and development programs for developing our ex vivo and in vivo cell engineering platforms, identifying and developing product candidates, and establishing manufacturing capabilities. Due to our early stage of development, the number of ongoing projects, and our ability to use resources across several projects, the majority of our research and development costs are not recorded on a program-specific basis. These include costs for personnel, laboratory, and other indirect facility and operating costs.
Research and development activities account for a significant portion of our operating expenses. We anticipate that our research and development expenses will increase for the foreseeable future as we expand our research and development efforts, including expanding the capabilities of our cell engineering platforms, identifying product candidates, completing existing preclinical studies and commencing new preclinical studies, commencing clinical trials, establishing internal and external manufacturing capabilities, seeking regulatory approval of our product candidates, and incurring costs to acquire and license technologies aligned with our goal of translating engineered cells to medicines. A change in the outcome of any of these factors could result in a significant change in the costs and timing associated with the development of our product candidates.
Research and development related success payments and contingent consideration
Research and development related success payments and contingent consideration include the change in the estimated fair value of our Cobalt Success Payment and Harvard Success Payment liabilities and Cobalt Contingent Consideration liability. Research and development expense related to our success payment liabilities and contingent consideration is unpredictable, including because our success payments are based, in part, on our common stock price and market capitalization at the end of each reporting period, and may continue to vary significantly from quarter to quarter and year to year due to changes in the assumptions used in the calculations.
General and administrative
General and administrative expenses consist of personnel-related costs, including salaries, benefits, and non-cash stock-based compensation for our employees in finance, legal, executive, human resources, information technology, and other administrative functions, legal and consulting fees, insurance fees, and facility costs not otherwise included in research and development expenses. Legal fees include those related to corporate and patent matters.
We anticipate that our general and administrative expenses will increase over the foreseeable future to support our continued research and development activities, grow our business, and support future possible business development opportunities. We also anticipate that we will continue to incur expenses related to audit and legal services associated with operating as a public company, maintaining compliance with the rules and regulations of the Securities and Exchange Commission (SEC) and standards applicable to companies listed on a national securities exchange, investor relations activities, and other administrative and professional services.
145
Results of operations
Comparison of the years ended December 31, 2021 and 2020
The following table summarizes our results of operations for the periods presented:
|
|
|
Year Ended December 31, |
|
|
|
|
|
|||||
|
|
|
2021 |
|
|
2020 |
|
|
Change |
|
|||
|
|
|
(in thousands) |
|
|||||||||
|
Operating expenses: |
|
|
|
|
|
|
|
|
|
|
|
|
|
Research and development |
|
$ |
248,626 |
|
|
$ |
132,944 |
|
|
$ |
115,682 |
|
|
Research and development related success payments and contingent consideration |
|
|
57,873 |
|
|
|
124,935 |
|
|
|
(67,062 |
) |
|
General and administrative |
|
|
50,410 |
|
|
|
28,270 |
|
|
|
22,140 |
|
|
Total operating expenses |
|
|
356,909 |
|
|
|
286,149 |
|
|
|
70,760 |
|
|
Loss from operations |
|
|
(356,909 |
) |
|
|
(286,149 |
) |
|
|
(70,760 |
) |
|
Interest income, net |
|
|
676 |
|
|
|
747 |
|
|
|
(71 |
) |
|
Other income, net |
|
|
305 |
|
|
|
97 |
|
|
|
208 |
|
|
Net loss |
|
$ |
(355,928 |
) |
|
$ |
(285,305 |
) |
|
$ |
(70,623 |
) |
Research and development expenses
The following table summarizes the components of our research and development expenses for the periods presented:
|
|
|
Year Ended December 31, |
|
|
|
|
|
|||||
|
|
|
2021 |
|
|
2020 |
|
|
Change |
|
|||
|
|
|
(in thousands) |
|
|||||||||
|
Acquisition and licensing of technology |
|
$ |
52,439 |
|
|
$ |
11,991 |
|
|
$ |
40,448 |
|
|
Personnel |
|
|
79,028 |
|
|
|
49,508 |
|
|
|
29,520 |
|
|
Research and laboratory |
|
|
59,908 |
|
|
|
31,913 |
|
|
|
27,995 |
|
|
Facility and other allocated costs |
|
|
47,730 |
|
|
|
30,215 |
|
|
|
17,515 |
|
|
Other |
|
|
9,521 |
|
|
|
9,317 |
|
|
|
204 |
|
|
Total research and development expense |
|
$ |
248,626 |
|
|
$ |
132,944 |
|
|
$ |
115,682 |
|
Research and development expense was $248.6 million and $132.9 million, respectively, for the years ended December 31, 2021 and 2020. The increase of $115.7 million was primarily due to:
|
|
• |
an increase in upfront license fees of $40.4 million due to the upfront expense of $50.0 million recorded in 2021 to license Beam’s genome editing technology, partially offset by the upfront expense of $8.5 million recorded in 2020 related to the acquisition of Oscine; |
|
|
• |
increased personnel-related expenses of $29.5 million, including non-cash stock-based compensation of $10.3 million, which was primarily attributable to an increase in headcount to expand our research and development capabilities; |
|
|
• |
an increase of $28.0 million in research and laboratory costs, including preclinical study, laboratory supply, third-party manufacturing, and other external research expenses; and |
|
|
• |
an increase of $17.5 million of facility and allocated costs, including rent, depreciation, and overhead. |
Research and development related success payments and contingent consideration
The following table summarizes the expenses associated with research and development related success payments and contingent consideration for the periods presented:
|
|
|
Year Ended December 31, |
|
|
|
|
|
|||||
|
|
|
2021 |
|
|
2020 |
|
|
Change |
|
|||
|
|
|
(in thousands) |
|
|||||||||
|
Success payments |
|
$ |
26,031 |
|
|
$ |
72,142 |
|
|
$ |
(46,111 |
) |
|
Contingent consideration |
|
|
31,842 |
|
|
|
52,793 |
|
|
|
(20,951 |
) |
|
Total research and development related success payments and contingent consideration |
|
$ |
57,873 |
|
|
$ |
124,935 |
|
|
$ |
(67,062 |
) |
For the years ended December 31, 2021 and 2020, we recognized non-cash expenses of $57.9 million and $124.9 million, respectively, for the changes in the estimated fair value of research and development related success payments and contingent
146
consideration. The expense related to the change in the estimated fair value of our Cobalt Success Payment and Harvard Success Payment liabilities in aggregate was $26.0 million and $72.1 million, respectively, for the years ended December 31, 2021 and 2020. The change in the estimated fair value of the success payment liabilities was due to a smaller increase in the growth of our market capitalization and common stock price during the relative periods. The expense related to the change in the estimated fair value of our Cobalt Contingent Consideration was $31.8 million and $52.8 million, respectively, for the years ended December 31, 2021 and 2020. The change in the estimated fair value of the Cobalt Contingent Consideration was primarily due to scientific progress toward the achievement of milestones during the relative periods.
General and administrative expenses
General and administrative expenses were $50.4 million and $28.3 million, respectively, for the years ended December 31, 2021 and 2020.The increase of $22.1 million was primarily due to increased personnel-related expenses of $10.8 million, including non-cash stock-based compensation of $6.2 million, primarily attributable to an increase in headcount to build our infrastructure and support our continued research and development activities, increased legal fees of $3.9 million to support our patent portfolio and licensing arrangements, increased insurance costs of $3.9 million associated with being a public company, increased consulting fees of $1.4 million, and increased facility costs, including rent, of $0.8 million.
Interest income, net
Interest income, net, was $0.7 million for each of the years ended December 31, 2021 and 2020 and consisted primarily of interest earned on our cash and marketable securities balances for each year.
Comparison of the years ended December 31, 2020 and 2019
The following table summarizes our results of operations for the periods presented:
|
|
|
Year Ended December 31, |
|
|
|
|
|
|||||
|
|
|
2020 |
|
|
2019 |
|
|
Change |
|
|||
|
|
|
(in thousands) |
|
|||||||||
|
Operating expenses: |
|
|
|
|
|
|
|
|
|
|
|
|
|
Research and development |
|
$ |
132,944 |
|
|
$ |
99,591 |
|
|
$ |
33,353 |
|
|
Research and development related success payments and contingent consideration |
|
|
124,935 |
|
|
|
19,784 |
|
|
|
105,151 |
|
|
General and administrative |
|
|
28,270 |
|
|
|
21,777 |
|
|
|
6,493 |
|
|
Total operating expenses |
|
|
286,149 |
|
|
|
141,152 |
|
|
|
144,997 |
|
|
Loss from operations |
|
|
(286,149 |
) |
|
|
(141,152 |
) |
|
|
(144,997 |
) |
|
Interest income, net |
|
|
747 |
|
|
|
2,856 |
|
|
|
(2,109 |
) |
|
Other income, net |
|
|
97 |
|
|
|
(29 |
) |
|
|
126 |
|
|
Loss before income taxes |
|
|
(285,305 |
) |
|
|
(138,325 |
) |
|
|
(146,980 |
) |
|
Benefit from income taxes |
|
|
- |
|
|
|
7,547 |
|
|
|
(7,547 |
) |
|
Net loss |
|
$ |
(285,305 |
) |
|
$ |
(130,778 |
) |
|
$ |
(154,527 |
) |
Research and development expenses
The following table summarizes the components of our research and development expenses for the periods presented:
|
|
|
Year Ended December 31, |
|
|
|
|
|
|||||||
|
|
|
2020 |
|
|
|
|
2019 |
|
|
Change |
|
|||
|
|
|
(in thousands) |
|
|||||||||||
|
Personnel |
|
$ |
49,508 |
|
|
|
|
$ |
30,378 |
|
|
$ |
19,130 |
|
|
Research and laboratory |
|
|
31,913 |
|
|
|
|
|
13,302 |
|
|
|
18,611 |
|
|
Facility and other allocated costs |
|
|
30,215 |
|
|
|
|
|
18,246 |
|
|
|
11,969 |
|
|
Acquisition and licensing of technology |
|
|
11,991 |
|
|
|
|
|
27,773 |
|
|
|
(15,782 |
) |
|
Other |
|
|
9,317 |
|
|
|
|
|
9,892 |
|
|
|
(575 |
) |
|
Total research and development expense |
|
$ |
132,944 |
|
|
|
|
$ |
99,591 |
|
|
$ |
33,353 |
|
Research and development expenses were $132.9 million and $99.6 million, respectively, for the years ended December 31, 2020 and 2019. The increase of $33.3 million was primarily due to:
|
|
• |
increased personnel-related expenses of $19.1 million, including non-cash stock-based compensation of $3.6 million, which was primarily attributable to an increase in headcount to expand our research and development capabilities; |
147
|
|
• |
an increase of $18.6 million in research and laboratory costs, including preclinical study, laboratory supply, and other external research expenses; and |
|
|
• |
an increase of $12.0 million of facility and other allocated costs, including rent, depreciation, and overhead. |
These increases were partially offset by a decline in costs to acquire and license technology of $15.8 million due to costs incurred in 2019 related to the Harvard and UCSF agreements, and the upfront fee for the acquisition of Cytocardia, partially offset by the upfront fee for the acquisition of Oscine in 2020.
Research and development related success payments and contingent consideration
The following table summarizes the expenses associated with research and development related success payments and contingent consideration for the periods presented:
|
|
|
Year Ended December 31, |
|
|
|
|
|
|||||
|
|
|
2020 |
|
|
2019 |
|
|
Change |
|
|||
|
|
|
(in thousands) |
|
|||||||||
|
Success payments |
|
$ |
72,142 |
|
|
$ |
1,924 |
|
|
$ |
70,218 |
|
|
Contingent consideration |
|
|
52,793 |
|
|
|
17,860 |
|
|
|
34,933 |
|
|
Total research and development related success payments and contingent consideration |
|
$ |
124,935 |
|
|
$ |
19,784 |
|
|
$ |
105,151 |
|
For the years ended December 31, 2020 and 2019, we recognized non-cash expenses of $124.9 million and $19.8 million, respectively, for the changes in the estimated fair value of research and development related success payments and contingent consideration. The expense related to the change in the estimated fair value of our Cobalt Success Payment and Harvard Success Payment liabilities in aggregate was $72.1 million and $1.9 million, respectively, for the years ended December 31, 2020 and 2019. The change in the estimated fair value of the success payment liabilities was due to increases in the estimated fair value of our Series A and Series B convertible preferred stock during the relative periods. The expense related to the change in the estimated fair value of our Cobalt Contingent Consideration was $52.8 million and $17.9 million, respectively, for the years ended December 31, 2020 and 2019. The change in the estimated fair value of the Cobalt Contingent Consideration was primarily due to scientific progress toward the achievement of milestones during the relative periods.
General and administrative expenses
General and administrative expenses were $28.3 million and $21.8 million, respectively, for the years ended December 31, 2020 and 2019. The increase of $6.5 million was primarily due to increased personnel-related expenses of $3.2 million, primarily attributable to an increase in headcount to build our infrastructure, increased information technology and facilities costs, including rent of $1.4 million, increased business taxes and insurance of $0.8 million, and increased consulting and legal fees of $0.5 million.
Interest income, net
Interest income, net was $0.7 million and $2.9 million, respectively, for the years ended December 31, 2020 and 2019. The decrease of $2.2 million was due to lower interest rates on cash and marketable securities balances.
Benefit from income taxes
The benefit from income taxes of $7.5 million for the year ended December 31, 2019 was due to a release of valuation allowance associated with the deferred tax liability of $7.5 million recorded in connection with the intangible asset from the Cobalt acquisition. There was no benefit from income taxes for the year ended December 31, 2020.
Liquidity, capital resources, and capital requirements
Sources of liquidity
As of December 31, 2021, we had $746.9 million in cash, cash equivalents, and marketable securities. To date we have raised an aggregate of approximately $1.3 billion in net proceeds through our IPO and private placements of our convertible preferred stock. Since our inception, we have not generated any revenue from product sales or any other sources, and we have incurred significant operating losses. We have not yet commercialized any products and we do not expect to generate revenue from sales of any product candidates for a number of years, if ever.
148
Future funding requirements
We expect to incur additional losses for the foreseeable future as we conduct and expand our research and development efforts, including conducting preclinical studies and clinical trials, developing new product candidates, establishing internal and external manufacturing capabilities, and funding our operations generally.
Based on our current operating plan, we believe that our existing cash, cash equivalents, and marketable securities will be sufficient to meet our working capital and capital expenditure needs for at least the next 12 months. However, we anticipate that we will need to raise additional financing in the future to fund our operations, including the commercialization of any approved product candidates. We are subject to the risks typically related to the development of new products, and we may encounter unforeseen expenses, difficulties, complications, delays, and other unknown factors that may adversely affect our business.
Our future capital requirements will depend on many factors, including:
|
|
• |
the scope, timing, progress, costs, and results of discovery, preclinical development, and clinical trials for our current or future product candidates; |
|
|
• |
the number and scope of clinical trials required for regulatory approval of our current or future product candidates; |
|
|
• |
the costs, timing, and outcome of regulatory review of our current or future product candidates; |
|
|
• |
the cost associated with building our manufacturing capabilities, as well as costs associated with the manufacturing of clinical and commercial supplies of our current and future product candidates; |
|
|
• |
the costs and timing of future commercialization activities, including manufacturing, marketing, sales, and distribution, for any of our product candidates for which we receive marketing approval; |
|
|
• |
the costs and timing of preparing, filing, and prosecuting patent applications, maintaining and enforcing our intellectual property rights, and defending any intellectual property-related claims, including any claims by third parties that we are infringing upon their intellectual property rights; |
|
|
• |
our ability to maintain existing, and establish new, strategic collaborations, licensing, or other arrangements and the financial terms of any such agreements, including the timing and amount of any future milestone, royalty, or other payments due under any such agreement; |
|
|
• |
the revenue, if any, received from commercial sales of our product candidates for which we receive marketing approval; |
|
|
• |
the expenses required to attract, hire, and retain skilled personnel; |
|
|
• |
the costs of operating as a public company; |
|
|
• |
our ability to establish a commercially viable pricing structure and obtain approval for coverage and adequate reimbursement from third-party and government payors; |
|
|
• |
potential interruptions or delays resulting from factors related to the ongoing COVID-19 pandemic; |
|
|
• |
the effect of competing technological and market developments; and |
|
|
• |
the extent to which we acquire or invest in businesses, products, and technologies. |
Until such time as we can generate significant revenue from product sales, if ever, we expect to finance our operations from the proceeds of equity or debt financings or capital obtained in connection with strategic collaborations or licensing or other arrangements. In the event that additional financing is required, we may not be able to raise it on terms that are acceptable to us or at all. If we raise additional funds through the issuance of equity or convertible debt securities, it may result in dilution to our existing stockholders. Debt financing, if available, may result in increased fixed payment obligations, and the existence of securities with rights that may be senior to those of our common stock. If we incur debt, we could become subject to covenants that would restrict our operations. If we raise funds through strategic collaborations or licensing or other arrangements, we may relinquish significant rights or grant licenses on terms that are not favorable to us. Our ability to raise additional funds may be adversely impacted by potential worsening global economic conditions and the recent disruptions to, and volatility in, the credit and financial markets in the United States and worldwide resulting from the ongoing COVID-19 pandemic or otherwise. If we are unable to raise additional capital when desired, our business, results of operations, and financial condition would be adversely affected.
149
Cash flows
The following table summarizes our cash flows for the periods indicated:
|
|
|
Year Ended December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
|
2019 |
|
|||
|
|
|
(in thousands) |
|
|||||||||
|
Net cash provided by (used in): |
|
|
|
|
|
|
|
|
|
|
|
|
|
Operating activities |
|
$ |
(251,054 |
) |
|
$ |
(137,982 |
) |
|
$ |
(85,518 |
) |
|
Investing activities |
|
|
(245,798 |
) |
|
|
(252,563 |
) |
|
|
(87,861 |
) |
|
Financing activities |
|
|
631,751 |
|
|
|
435,687 |
|
|
|
223,740 |
|
|
Net increase in cash, cash equivalents, and restricted cash |
|
$ |
134,899 |
|
|
$ |
45,142 |
|
|
$ |
50,361 |
|
Operating activities
During the year ended December 31, 2021, net cash used in operating activities was $251.0 million, consisting primarily of our net loss of $355.9 million, partially offset by the change in our net operating assets and liabilities of $9.8 million and non-cash charges of $95.1 million. The non-cash charges of $95.1 million consisted of $31.8 million for revaluation of contingent consideration, $26.0 million for revaluation of our success payment liabilities, non-cash stock-based compensation expense of $22.4 million, depreciation expense of $11.1 million, and other non-cash charges of $3.8 million.
During the year ended December 31, 2020, net cash used in operating activities was $138.0 million, consisting primarily of our net loss of $285.3 million partially offset by non-cash charges of $141.2 million and an increase in our net operating assets of $6.2 million. The non-cash charges of $141.2 million consisted of $72.1 million for revaluation of our success payment liabilities, $52.8 million for revaluation of contingent consideration, depreciation expense of $5.9 million, non-cash stock-based compensation expense of $5.8 million, and other non-cash charges of $4.6 million.
During the year ended December 31, 2019, net cash used in operating activities was $85.5 million, consisting primarily of our net loss of $130.8 million and a tax benefit of $7.5 million recorded in connection with the Cobalt acquisition, partially offset by non-cash charges of $42.3 million and an increase in our net operating assets of $10.5 million. The non-cash charges of $42.3 million consisted of $17.9 million for revaluation of contingent consideration, $11.9 million for the issuance of stock in connection with license agreements, $6.5 million for revaluation of success payment and contingent liabilities, depreciation expense of $1.8 million, and other non-cash charges of $4.2 million.
Investing activities
During the years ended December 31, 2021, 2020 and 2019, cash used in investing activities was $245.8 million, $252.6 million, and $87.9 million, respectively. This consisted primarily of net purchases, sales, and maturities of marketable securities of $211.3 million, $228.7 million and $58.5 million, respectively, and purchases of property and equipment of $29.9 million, $23.9 million and $26.2 million, respectively, for the years ended December 31, 2021, 2020 and 2019.
Financing activities
During the year ended December 31, 2021, cash provided by financing activities was $631.7 million, consisting primarily of net proceeds from our IPO of $626.4 million and $5.3 million in proceeds from our employee stock purchase plan and the exercise of stock options.
During the years ended December 31, 2020 and 2019, cash provided by financing activities was $435.7 million and $223.7 million, respectively, consisting primarily of net proceeds from the sale of our convertible preferred stock.
Contractual obligations and commitments
The following table summarizes our significant contractual obligations and commitments as of December 31, 2021:
|
|
|
Payments Due by Period |
|
|||||||||||||||||
|
|
|
Less than 1 Year |
|
|
1 to 3 Years |
|
|
3 to 5 Years |
|
|
More than 5 Years |
|
|
Total |
|
|||||
|
|
|
(in thousands) |
|
|||||||||||||||||
|
Operating lease obligations |
|
$ |
19,954 |
|
|
$ |
42,112 |
|
|
$ |
38,150 |
|
|
$ |
80,913 |
|
|
$ |
181,129 |
|
150
Other than as disclosed in the table above, the payment obligations under our license, collaboration, and acquisition agreements as of December 31, 2021 are contingent upon future events such as our achievement of pre-specified development, regulatory, and commercial milestones or royalties on net product sales. See the section titled “Business—Key Intellectual Property Agreements” for more information about these payment obligations.
We are also obligated to make a success payment to Cobalt of up to $500.0 million, payable in cash or stock, pursuant to the terms and conditions in the Cobalt acquisition agreement, and up to an aggregate of $175.0 million in success payments to Harvard, payable in cash. See the subsection below titled “—Critical accounting policies and significant judgments and estimates—Success payments” and Note 3, Acquisitions, and Note 4, License and collaboration agreements, to our consolidated financial statements located elsewhere in this Annual Report for more information on the success payments. As of December 31, 2021, the timing and likelihood of achieving the milestones and success payments and generating future product sales are uncertain, and therefore any related payments are not included in the table above.
We also enter into agreements in the normal course of business for sponsored research, preclinical studies, contract manufacturing, and other services and products for operating purposes, which are generally cancelable upon written notice. These obligations and commitments are not included in the table above.
Off-balance sheet arrangements
Since our inception, we have not engaged in any off-balance sheet arrangements as defined under the rules and regulations of the SEC.
JOBS Act accounting election
We are an “emerging growth company,” as defined in the Jumpstart Our Business Startups Act of 2012 (the JOBS Act). We will cease to be an emerging growth company until the earliest of (1) December 31, 2026, (2) the last day of the fiscal year in which we have total annual gross revenue of at least $1.07 billion, (3) the last day of the fiscal year in which we are deemed to be a “large accelerated filer” as defined in Rule 12b-2 under the Exchange Act, which would occur if the fair market value of our common stock held by non-affiliates exceeded $700.0 million as of the last business day of the second fiscal quarter of such year, or (4) the date on which we have issued more than $1.0 billion in non-convertible debt securities during the prior three-year period. For as long as we remain an emerging growth company, we are permitted and intend to rely on certain exemptions from various public company reporting requirements, including not being required to have our independent registered public accounting firm provide an attestation report on our internal control over financial reporting pursuant to Section 404(b) of the Sarbanes-Oxley Act of 2002, reduced disclosure obligations regarding executive compensation in our periodic reports and proxy statements, and exemptions from the requirements of holding a nonbinding advisory vote on executive compensation and any golden parachute payments not previously approved. In addition, under the JOBS Act, emerging growth companies can delay adopting new or revised accounting standards issued subsequent to the enactment of the JOBS Act until such time as those standards apply to private companies. We have elected to use the extended transition period for any new or revised accounting standards during the period in which we remain an emerging growth company; however, we may adopt certain new or revised accounting standards early if the standard allows early adoption.
Critical accounting policies and significant judgments and estimates
Our consolidated financial statements are prepared in accordance with generally accepted accounting principles in the United States. The preparation of these financial statements requires us to make estimates and assumptions that affect the amounts reported in the consolidated financial statements and accompanying notes. Our estimates are based on our historical experience and on various other factors that we believe are reasonable under the circumstances, the results of which form the basis for making judgments about the carrying value of assets and liabilities that are not readily apparent from other sources. Actual results may differ from these estimates under different assumptions or conditions. Our significant accounting policies are described in more detail in the notes to our consolidated financial statements included elsewhere in this Annual Report. We believe the following accounting policies relate to the significant areas involving management’s judgments and estimates and are critical to understanding our historical and future performance.
Research and development expenses
We record research and development costs in the periods in which they are incurred. We accrue for research and development costs based on the estimated services performed, but not yet invoiced, pursuant to contracts with research institutions or other service providers that conduct and manage preclinical studies and other research services on our behalf and record these costs in accrued and other current liabilities. We make judgments and estimates in determining the accrued liabilities balance at each reporting period. Payments made prior to the receipt of goods or services to be used in research and development are recorded as prepaid expenses until the goods or services are received.
151
To date, we have not experienced any material differences between accrued costs and actual costs incurred. However, the status and timing of actual services performed may vary from our estimates, resulting in adjustments to expense in future periods. Changes in these estimates that result in material changes to our accruals could materially affect our results of operations.
Acquisitions
We account for business combinations using the acquisition method of accounting, which requires the assets acquired, including in-process research and development (IPR&D), and liabilities assumed, be recorded at their fair values as of the acquisition date. Any excess of the purchase price over the fair value of net assets acquired is recorded as goodwill. The determination of the estimated fair value of these items requires us to make significant estimates and assumptions.
If we determine the acquisition does not meet the definition of a business combination under the acquisition method of accounting, the transaction is accounted for as an asset acquisition and no goodwill or contingent consideration are recognized at the acquisition date. In an asset acquisition, up-front payments allocated to IPR&D are recorded in research and development expense if it is determined that there is no alternative future use, and subsequent milestone payments are recorded in research and development expense when achieved.
Intangible assets and goodwill
Accounting for business combinations requires us to make significant estimates and assumptions with respect to tangible and intangible assets acquired and liabilities assumed. We use our best estimates and assumptions to accurately assign fair value to the tangible and intangible assets acquired and liabilities assumed at the acquisition date as well as the useful lives of those acquired intangible assets. Intangible assets are reviewed for impairment annually and upon the occurrence of triggering events or substantive changes in circumstances that could indicate a potential impairment.
Goodwill represents the excess of the purchase price over the estimated fair value of the identifiable assets acquired and liabilities assumed in a business combination. We evaluate goodwill for impairment annually and upon the occurrence of triggering events or substantive changes in circumstances that could indicate a potential impairment. Our evaluation includes assessing qualitative factors or performing a quantitative analysis to determine whether it is more-likely-than-not that the fair value of net assets is below the carrying amounts.
Contingent consideration
Contingent consideration obligations are estimated at fair value at the acquisition date of a business combination and at each subsequent balance sheet date, with changes in fair value recorded in research and development related success payments and contingent consideration. The fair value of contingent consideration is determined by calculating the probability-weighted estimated value of the milestone payments based on the assessment of the likelihood and estimated timing that the milestones would be achieved and applying the relevant discount rates. We utilize significant estimates and assumptions in determining the estimated contingent consideration and associated expense or gain at each balance sheet date. The valuation of contingent consideration uses assumptions we believe would be made by a market participant. In evaluating the fair value of contingent consideration, a significant amount of judgment is required to estimate the likelihood and timing that the milestones would be achieved. We assess these estimates on an on-going basis as additional data impacting the assumptions become available. Contingent consideration may change significantly as development progresses and additional data is obtained, impacting our assumptions regarding probabilities of successful achievement of the related milestones used to estimate the fair value of the liability and the timing in which they are expected to be achieved. Accordingly, the use of different market assumptions and/or different valuation techniques could result in materially different fair value estimates.
Success payments
The Cobalt Success Payment was recorded as a liability on the consolidated balance sheet at fair value on the acquisition date and is remeasured at each subsequent reporting period, with changes in fair value recognized in research and development related success payments and contingent consideration. For the Harvard Success Payments, both the initial value and subsequent changes in fair value are recorded in research and development related success payments and contingent consideration. To determine the estimated fair value of the success payment liabilities, we use a Monte Carlo simulation methodology which models the estimated fair value of the liability based on several key assumptions, including the estimated number and timing of valuation measurement dates on the basis of which payments may be triggered, term of the success payments, the risk-free interest rate, and expected volatility, which is estimated using peer company stocks for a period of time commensurate with the expected term assumption. Prior to our IPO, the calculation of the Harvard Success Payment liability incorporated the estimated fair value of our Series A convertible preferred stock, and the Cobalt Success Payment liability incorporated our estimated future value implied by the per share value of the Series B convertible preferred stock at issuance, or any security into which such stock had been converted or exchanged. Concurrent with the
152
closing of our IPO in February 2021, the Series A and Series B convertible preferred stock converted into common stock. Subsequent to our IPO, the computation of the estimated fair value of the Harvard Success Payments incorporates the per share fair market value of our common stock at the end of each reporting period, and the computation of the estimated fair value of the Cobalt Success Payment incorporates our market capitalization at the end of each reporting period. The assumptions used to calculate the fair value of the success payments are subject to a significant amount of judgment and a small change in the assumptions may have a relatively large change in the estimated liability and resulting expense or gain.
Stock-based compensation
We recognize compensation costs related to restricted stock awards, restricted stock units, and stock options granted to employees and nonemployees based on the estimated fair value of the awards on the date of grant, and we recognize forfeitures as they occur. For restricted stock awards and restricted stock units, the fair value of our common stock is used to determine the resulting stock-based compensation expense. For stock options, we estimate the grant date fair value, and the resulting stock-based compensation expense, using the Black-Scholes option pricing model. The fair value of the stock-based awards is recognized as an expense on a straight-line basis over the requisite service period, which is generally the vesting period.
The Black-Scholes option pricing model requires the use of highly subjective assumptions to determine the fair value of stock-based awards. These assumptions include:
|
|
• |
Fair Value of Common Stock—For all periods prior to our IPO, there was no active public market for our common stock. Therefore, our board of directors, with the assistance and upon the recommendation of management, had, for financial reporting purposes, periodically determined the estimated per share fair value of our common stock on the date of grant using contemporaneous independent third-party valuations consistent with the American Institute of Certified Public Accountants Practice Aid, Valuation of Privately-Held Company Equity Securities Issued as Compensation, as well as a range of factors and assumptions. The determination of such estimates involved inherent uncertainties, and our board of directors exercised significant judgment in estimating the value of our common stock. As a result, if factors or expected outcomes changed or we had used significantly different assumptions or estimates, our equity-based compensation could have been materially different. Following the closing of our IPO, the fair value of common stock is based on the closing price as reported on The Nasdaq Global Select Market on the date of grant. |
|
|
• |
Expected Term—The expected term represents the period that the stock-based awards are expected to be outstanding. We use the simplified method to determine the expected term, which is based on the average of the time-to-vesting and the contractual life of the options. |
|
|
• |
Expected Volatility—Due to our limited operating history and lack of company-specific historical and implied volatility data, the expected volatility is estimated based on the average historical volatilities of common stock of comparable publicly traded entities over a period of time commensurate with the expected term of the stock option grants. The comparable companies are chosen based on their size, stage in the product development cycle, or area of specialty. We will continue to apply this process until sufficient historical information regarding the volatility of our own stock price becomes available. |
|
|
• |
Risk-Free Interest Rate—The risk-free interest rate is based on the U.S. Treasury yield in effect at the time of grant for zero-coupon U.S. Treasury notes with maturities approximately equal to the expected term of the awards. |
|
|
• |
Expected Dividend—We have never paid dividends on our common stock and have no plans to pay dividends on our common stock. Therefore, we used an expected dividend yield of zero. |
See Note 12, Stock-based compensation to our consolidated financial statements included elsewhere in this Annual Report for information concerning certain specific assumptions we used in applying the Black-Scholes option pricing model to determine the estimated fair value of our stock options granted in the years ended December 31, 2021, 2020, and 2019. Such assumptions involve inherent uncertainties and the application of significant judgment. As a result, if factors or expected outcomes change or we use significantly different assumptions or estimates, our stock-based compensation could be materially different.
Recently adopted and recent accounting pronouncements
See Note 2, Summary of significant accounting policies to our consolidated financial statements included elsewhere in this Annual Report for information about recent accounting pronouncements, the timing of their adoption, and our assessment, to the extent we have made one yet, of their potential impact on our financial condition or results of operations.
153
Item 7A. Quantitative and Qualitative Disclosures About Market Risk.
We are exposed to market risks in the ordinary course of our business primarily related to interest rate sensitivities and the volatility of our common stock price.
Interest rate risk
As of December 31, 2021, we had cash, cash equivalents, and restricted cash of $261.8 million, which consisted of bank deposits and money market funds, and also had marketable securities of $493.8 million. The primary objective of our investment activities is to preserve capital to fund our operations while earning a low risk return. Because our marketable securities are primarily short-term in duration, we believe that our exposure to interest rate risk is not significant, and a hypothetical 10% change in market interest rates during any of the periods presented would not have had a significant impact on the total value of our portfolio. We had no debt outstanding as of December 31, 2021.
Market capitalization and common stock price sensitivity
We agreed to make a success payment to Cobalt based on our market capitalization payable in cash or stock, and success payments to Harvard based on increases in the per share fair market value of our common stock, payable in cash.
As of December 31, 2021, the estimated fair value of the success payment liabilities was $102.5 million. For the twelve months ended December 31, 2021, we recorded an expense of $26.0 million related to the aggregate change in the estimated fair value of our success payment liabilities.
Changes in our market capitalization and the fair value of our common stock as of each balance date may have a relatively large change in the estimated valuation of the success payment liabilities and resulting expense or gain. See Item 1A. Risk Factors included in this Annual Report for a sensitivity analysis showing the impact that a hypothetical change in our market capitalization and common stock value would have had on our results for the year ended December 31, 2021.
Foreign currency sensitivity
We are not currently exposed to significant market risk related to changes in foreign currency exchange rates; however, we do contract with vendors that are located outside of the United States and may be subject to fluctuations in foreign currency rates. We may enter into additional contracts with vendors located outside of the United States in the future, which may increase our foreign currency exchange risk.
154
Item 8. Financial Statements and Supplementary Data.
SANA BIOTECHNOLOGY, INC.
INDEX TO CONSOLIDATED FINANCIAL STATEMENTS
|
(PCAOB ID: |
156 |
|
|
157 |
||
|
158 |
||
|
159 |
||
|
Consolidated Statements of Convertible Preferred Stock and Stockholders’ Equity (Deficit) |
160 |
|
|
161 |
||
|
162 |
||
155
Report of Independent Registered Public Accounting Firm
To the Stockholders and the Board of Directors of Sana Biotechnology, Inc.
Opinion on the Financial Statements
We have audited the accompanying consolidated balance sheets of Sana Biotechnology Inc. (the Company) as of December 31, 2021 and 2020, the related consolidated statements of operations, comprehensive loss, convertible preferred stock and stockholders’ equity (deficit) and cash flows for each of the three years in the period ended December 31, 2021, and the related notes (collectively referred to as the “consolidated financial statements”). In our opinion, the consolidated financial statements present fairly, in all material respects, the financial position of the Company at December 31, 2021 and 2020, and the results of its operations and its cash flows for each of the three years in the period ended December 31, 2021, in conformity with U.S. generally accepted accounting principles.
Basis for Opinion
These financial statements are the responsibility of the Company’s management. Our responsibility is to express an opinion on the Company’s financial statements based on our audits. We are a public accounting firm registered with the Public Company Accounting Oversight Board (United States) (PCAOB) and are required to be independent with respect to the Company in accordance with the U.S. federal securities laws and the applicable rules and regulations of the Securities and Exchange Commission and the PCAOB.
We conducted our audits in accordance with the standards of the PCAOB. Those standards require that we plan and perform the audit to obtain reasonable assurance about whether the financial statements are free of material misstatement, whether due to error or fraud. The Company is not required to have, nor were we engaged to perform, an audit of its internal control over financial reporting. As part of our audits we are required to obtain an understanding of internal control over financial reporting but not for the purpose of expressing an opinion on the effectiveness of the Company’s internal control over financial reporting. Accordingly, we express no such opinion.
Our audits included performing procedures to assess the risks of material misstatement of the financial statements, whether due to error or fraud, and performing procedures that respond to those risks. Such procedures included examining, on a test basis, evidence regarding the amounts and disclosures in the financial statements. Our audits also included evaluating the accounting principles used and significant estimates made by management, as well as evaluating the overall presentation of the financial statements. We believe that our audits provide a reasonable basis for our opinion.
/s/
We have served as the Company’s auditor since 2018.
March 16, 2022
156
Sana Biotechnology, Inc.
Consolidated Balance Sheets
(in thousands, except per share amounts)
|
|
|
December 31, |
|
|||||
|
|
|
2021 |
|
|
2020 |
|
||
|
ASSETS |
|
|
|
|
|
|
|
|
|
Current assets: |
|
|
|
|
|
|
|
|
|
Cash and cash equivalents |
|
$ |
|
|
|
$ |
|
|
|
Marketable securities |
|
|
|
|
|
|
|
|
|
Prepaid expenses and other current assets |
|
|
|
|
|
|
|
|
|
Total current assets |
|
|
|
|
|
|
|
|
|
Long-term marketable securities |
|
|
|
|
|
|
|
|
|
Property and equipment, net |
|
|
|
|
|
|
|
|
|
Operating lease right-of-use assets |
|
|
|
|
|
|
|
|
|
Restricted cash |
|
|
|
|
|
|
|
|
|
Intangible asset |
|
|
|
|
|
|
|
|
|
Goodwill |
|
|
|
|
|
|
|
|
|
Other non-current assets |
|
|
|
|
|
|
|
|
|
TOTAL ASSETS |
|
$ |
|
|
|
$ |
|
|
|
LIABILITIES, CONVERTIBLE PREFERRED STOCK, AND STOCKHOLDERS' EQUITY (DEFICIT) |
|
|
|
|
|
|
|
|
|
Current liabilities: |
|
|
|
|
|
|
|
|
|
Accounts payable |
|
$ |
|
|
|
$ |
|
|
|
Accrued compensation |
|
|
|
|
|
|
|
|
|
Accrued expenses and other current liabilities |
|
|
|
|
|
|
|
|
|
Operating lease liabilities |
|
|
|
|
|
|
|
|
|
Contingent consideration |
|
|
|
|
|
|
- |
|
|
Success payment liabilities |
|
|
|
|
|
|
- |
|
|
Total current liabilities |
|
|
|
|
|
|
|
|
|
Operating lease liabilities, net of current portion |
|
|
|
|
|
|
|
|
|
Contingent consideration, net of current portion |
|
|
|
|
|
|
|
|
|
Success payment liabilities, net of current portion |
|
|
|
|
|
|
|
|
|
Other non-current liabilities |
|
|
- |
|
|
|
|
|
|
Total liabilities |
|
|
|
|
|
|
|
|
|
Commitments and contingencies (Note 9) |
|
|
|
|
|
|
|
|
|
Convertible preferred stock, $ as of December 31, 2021 and 2020, respectively; outstanding as of December 31, 2021 and 2020, respectively |
|
|
- |
|
|
|
|
|
|
Stockholders' equity (deficit): |
|
|
|
|
|
|
|
|
|
Preferred stock, $ December 31, 2021 and 2020, respectively; December 31, 2021 and 2020 |
|
|
|
|
|
|
|
|
|
Common stock, $ December 31, 2021 and 2020, respectively; outstanding as of December 31, 2021 and 2020, respectively |
|
|
|
|
|
|
|
|
|
Additional paid-in capital |
|
|
|
|
|
|
|
|
|
Accumulated other comprehensive income (loss) |
|
|
( |
) |
|
|
|
|
|
Accumulated deficit |
|
|
( |
) |
|
|
( |
) |
|
Total stockholders' equity (deficit) |
|
|
|
|
|
|
( |
) |
|
TOTAL LIABILITIES, CONVERTIBLE PREFERRED STOCK, AND STOCKHOLDERS' EQUITY (DEFICIT) |
|
$ |
|
|
|
$ |
|
|
The accompanying notes are an integral part of these consolidated financial statements.
157
Sana Biotechnology, Inc.
Consolidated Statements of Operations
(in thousands, except per share amounts)
|
|
|
Year Ended December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
|
2019 |
|
|||
|
Operating expenses: |
|
|
|
|
|
|
|
|
|
|
|
|
|
Research and development |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
Research and development related success payments and contingent consideration |
|
|
|
|
|
|
|
|
|
|
|
|
|
General and administrative |
|
|
|
|
|
|
|
|
|
|
|
|
|
Total operating expenses |
|
|
|
|
|
|
|
|
|
|
|
|
|
Loss from operations |
|
|
( |
) |
|
|
( |
) |
|
|
( |
) |
|
Interest income, net |
|
|
|
|
|
|
|
|
|
|
|
|
|
Other income (expense), net |
|
|
|
|
|
|
|
|
|
|
( |
) |
|
Loss before income taxes |
|
|
( |
) |
|
|
( |
) |
|
|
( |
) |
|
Benefit from income taxes |
|
|
— |
|
|
|
— |
|
|
|
|
|
|
Net loss |
|
$ |
( |
) |
|
$ |
( |
) |
|
$ |
( |
) |
|
Net loss per common share - basic and diluted |
|
$ |
( |
) |
|
$ |
( |
) |
|
$ |
( |
) |
|
Weighted-average number of common shares - basic and diluted |
|
|
|
|
|
|
|
|
|
|
|
|
The accompanying notes are an integral part of these consolidated financial statements.
158
Sana Biotechnology, Inc.
Consolidated Statements of Comprehensive Loss
(in thousands)
|
|
|
Year Ended December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
|
2019 |
|
|||
|
Net loss |
|
$ |
( |
) |
|
$ |
( |
) |
|
$ |
( |
) |
|
Other comprehensive income (loss), net of tax: |
|
|
|
|
|
|
|
|
|
|
|
|
|
Unrealized gain (loss) on marketable securities, net |
|
|
( |
) |
|
|
|
|
|
|
|
|
|
Total comprehensive loss |
|
$ |
( |
) |
|
$ |
( |
) |
|
$ |
( |
) |
The accompanying notes are an integral part of these consolidated financial statements.
159
Sana Biotechnology, Inc.
Consolidated Statements of Convertible Preferred Stock and Stockholders’ Equity (Deficit)
(in thousands)
|
|
|
Convertible Preferred Stock |
|
|
|
Common Stock |
|
|
Additional Paid-In |
|
|
Accumulated Other Comprehensive |
|
|
Accumulated |
|
|
Total Stockholders' |
|
||||||||||||||
|
|
|
Shares |
|
|
Amount |
|
|
|
Shares |
|
|
Amount |
|
|
Capital |
|
|
Income (Loss) |
|
|
Deficit |
|
|
Equity (Deficit) |
|
||||||||
|
Balance as of December 31, 2018 |
|
|
|
|
|
$ |
|
|
|
|
|
|
|
|
$ |
- |
|
|
$ |
|
|
|
$ |
- |
|
|
$ |
( |
) |
|
$ |
( |
) |
|
Adjustment to beginning accumulated deficit from adoption of ASC 842 |
|
|
- |
|
|
|
- |
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
( |
) |
|
|
( |
) |
|
Issuance of Series A-2 convertible preferred stock, net of $ |
|
|
|
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
Issuance of Series A-2 convertible preferred stock for acquisition, non-cash |
|
|
|
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
Issuance of Series A-2 convertible preferred stock in connection with license agreements |
|
|
|
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
Vesting of restricted stock |
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Stock-based compensation |
|
|
- |
|
|
|
- |
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Exercise of stock options |
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
|
- |
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Unrealized gain on marketable securities, net |
|
|
- |
|
|
|
- |
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
- |
|
|
|
|
|
|
Net loss |
|
|
- |
|
|
|
- |
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
( |
) |
|
|
( |
) |
|
Balance as of December 31, 2019 |
|
|
|
|
|
$ |
|
|
|
|
|
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
( |
) |
|
$ |
( |
) |
|
Issuance of Series B convertible preferred stock, net of $ |
|
|
|
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
Issuance of common stock in connection with license agreements |
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
|
- |
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Vesting of restricted stock |
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Stock-based compensation |
|
|
- |
|
|
|
- |
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Exercise of stock options |
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
|
- |
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Unrealized gain on marketable securities, net |
|
|
- |
|
|
|
- |
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
- |
|
|
|
|
|
|
Net loss |
|
|
- |
|
|
|
- |
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
( |
) |
|
|
( |
) |
|
Balance as of December 31, 2020 |
|
|
|
|
|
$ |
|
|
|
|
|
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
( |
) |
|
$ |
( |
) |
|
Conversion of convertible preferred stock into common stock upon initial public offering |
|
|
( |
) |
|
|
( |
) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Issuance of common stock in initial public offering, net of $ |
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Vesting of restricted stock |
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
Exercise of stock options |
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
|
- |
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Issuance of common stock related to employee stock purchase plan |
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
|
- |
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Stock-based compensation |
|
|
- |
|
|
|
- |
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Unrealized loss on marketable securities, net |
|
|
- |
|
|
|
- |
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
( |
) |
|
|
- |
|
|
|
( |
) |
|
Net loss |
|
|
- |
|
|
|
- |
|
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
- |
|
|
|
( |
) |
|
|
( |
) |
|
Balance as of December 31, 2021 |
|
|
- |
|
|
$ |
- |
|
|
|
|
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
( |
) |
|
$ |
( |
) |
|
$ |
|
|
The accompanying notes are an integral part of these consolidated financial statements.
160
Sana Biotechnology, Inc.
Consolidated Statements of Cash Flows
(in thousands)
|
|
|
Year Ended December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
|
2019 |
|
|||
|
OPERATING ACTIVITIES: |
|
|
|
|
|
|
|
|
|
|
|
|
|
Net loss |
|
$ |
( |
) |
|
$ |
( |
) |
|
$ |
( |
) |
|
Adjustments to reconcile net loss to net cash used in operating activities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
Depreciation |
|
|
|
|
|
|
|
|
|
|
|
|
|
Deferred income tax |
|
|
- |
|
|
|
- |
|
|
|
( |
) |
|
Stock-based compensation expense |
|
|
|
|
|
|
|
|
|
|
|
|
|
Change in the estimated fair value of contingent consideration |
|
|
|
|
|
|
|
|
|
|
|
|
|
Change in the estimated fair value of success payment liabilities |
|
|
|
|
|
|
|
|
|
|
|
|
|
Non-cash expense in connection with license agreement and asset acquisition |
|
|
- |
|
|
|
|
|
|
|
|
|
|
Non-cash expense for operating lease right-of-use assets |
|
|
|
|
|
|
|
|
|
|
|
|
|
Other non-cash items, net |
|
|
( |
) |
|
|
( |
) |
|
|
( |
) |
|
Changes in operating assets and liabilities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
Prepaid expenses and other assets |
|
|
( |
) |
|
|
( |
) |
|
|
( |
) |
|
Operating lease right-of-use assets and liabilities |
|
|
|
|
|
|
|
|
|
|
|
|
|
Accounts payable |
|
|
( |
) |
|
|
|
|
|
|
( |
) |
|
Accrued expenses and other liabilities |
|
|
|
|
|
|
|
|
|
|
|
|
|
Net cash used in operating activities |
|
|
( |
) |
|
|
( |
) |
|
|
( |
) |
|
INVESTING ACTIVITIES: |
|
|
|
|
|
|
|
|
|
|
|
|
|
Purchases of marketable securities |
|
|
( |
) |
|
|
( |
) |
|
|
( |
) |
|
Proceeds from sales and maturities of marketable securities |
|
|
|
|
|
|
|
|
|
|
|
|
|
Purchases of property and equipment |
|
|
( |
) |
|
|
( |
) |
|
|
( |
) |
|
Acquisitions, net of cash acquired |
|
|
- |
|
|
|
- |
|
|
|
( |
) |
|
Other investing activities |
|
|
( |
) |
|
|
- |
|
|
|
- |
|
|
Net cash used in investing activities |
|
|
( |
) |
|
|
( |
) |
|
|
( |
) |
|
FINANCING ACTIVITIES: |
|
|
|
|
|
|
|
|
|
|
|
|
|
Proceeds from initial public offering, net of issuance costs |
|
|
|
|
|
|
- |
|
|
|
- |
|
|
Proceeds from issuance of convertible preferred stock, net of issuance costs |
|
|
- |
|
|
|
|
|
|
|
|
|
|
Proceeds from employee stock purchase plan and exercise of stock options, net |
|
|
|
|
|
|
|
|
|
|
|
|
|
Net cash provided by financing activities |
|
|
|
|
|
|
|
|
|
|
|
|
|
Net increase in cash, cash equivalents, and restricted cash |
|
|
|
|
|
|
|
|
|
|
|
|
|
Cash, cash equivalents, and restricted cash at beginning of period |
|
|
|
|
|
|
|
|
|
|
|
|
|
Cash, cash equivalents, and restricted cash at end of period |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
SUPPLEMENTAL CASH FLOW INFORMATION: |
|
|
|
|
|
|
|
|
|
|
|
|
|
Operating lease right-of-use assets obtained in exchange for lease obligations |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
Cash received for amounts related to tenant improvement allowances |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
Purchases of property and equipment included in accounts payable and accrued liabilities |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
Issuance of convertible preferred stock for acquisition |
|
$ |
- |
|
|
$ |
- |
|
|
$ |
|
|
The accompanying notes are an integral part of these consolidated financial statements.
161
Sana Biotechnology, Inc.
Notes to Consolidated Financial Statements
1. Organization
Sana Biotechnology, Inc. (the Company or Sana) is a biotechnology company focusing on utilizing engineered cells as medicines. The Company’s operations to date have included identifying and developing potential product candidates, executing preclinical studies, establishing manufacturing capabilities, acquiring technology, organizing and staffing the Company, business planning, establishing the Company’s intellectual property portfolio, raising capital, and providing general and administrative support for these operations.
Reverse stock split
Initial public offering
In February 2021, the Company successfully completed its initial public offering (IPO) of its common stock. In connection with its IPO, the Company issued
Need for additional capital
The Company is subject to a number of risks and uncertainties similar to other biotechnology companies in the development stage, including, but not limited to, those related to the need to obtain adequate additional funding, possible failure of preclinical testing or clinical trials, the need to obtain marketing approval for its product candidates, building out internal and external manufacturing capabilities, competitors developing new technological innovations, the need to successfully commercialize and gain market acceptance of the Company’s products, the need to protect the Company’s intellectual property and proprietary technology, and the need to attract and retain key scientific and management personnel. If the Company does not successfully commercialize or partner any of its product candidates, it will be unable to generate product revenue or achieve profitability. Until such time as the Company can generate significant revenue from product sales, if ever, it expects to finance its operations with the proceeds from additional equity or debt financings or capital obtained in connection with strategic collaborations or licensing or other arrangements. In the event that additional financing is required, the Company may not be able to raise it on terms acceptable to it or at all.
The Company has incurred operating losses each year since inception and expects such losses to continue for the foreseeable future. As of December 31, 2021, the Company had cash, cash equivalents, and marketable securities of $
2. Summary of significant accounting policies
Basis of presentation
The accompanying consolidated financial statements include the accounts of the Company and its wholly-owned subsidiaries. The Company’s consolidated financial statements have been prepared in accordance with generally accepted accounting principles in the United States (GAAP). Certain prior period amounts have been reclassified to conform to current period presentation.
162
Use of estimates
The preparation of the financial statements in conformity with GAAP requires management to make estimates and assumptions that affect the amounts reported in the financial statements and accompanying notes. The Company evaluates its estimates and assumptions on an ongoing basis using historical experience and other factors and adjusts those estimates and assumptions when facts and circumstances dictate. Actual results could materially differ from those estimates. The most significant estimates in the Company’s consolidated financial statements relate to success payment liabilities, contingent consideration, business combinations, accrued expenses, operating lease right of use assets and liabilities, and the valuation of stock options.
Cash and cash equivalents
Cash and cash equivalents include cash and highly liquid investments with original maturities of three months or less at acquisition. Cash equivalents include investments in money market funds with commercial banks and financial institutions and are stated at fair value.
Marketable securities
Marketable securities are classified as available-for-sale debt securities and are carried at fair value, which is derived from independent pricing sources based on quoted prices in active markets for similar securities. Investments in securities with maturities of less than one year, or those for which management intends to use to fund current operations, are included in current assets. Unrealized gains and losses that are deemed to be temporary in nature are reported as a component of accumulated comprehensive income (loss). Amortization, accretion, and dividends are included in other income (expense), net on the consolidated statement of operations. The cost of securities sold is based on the specific-identification method. Each reporting period, the Company evaluates whether declines in fair value below carrying value are due to expected credit losses, as well as the Company’s ability and intent to hold the investment until a forecasted recovery occurs. Expected credit losses are recorded as an allowance through other income (expense), net.
Concentrations of credit risk and off-balance sheet risk
The Company maintains its cash, cash equivalents, and marketable securities with high quality, accredited financial institutions. These amounts, at times, may exceed federally insured limits. The Company has not experienced any credit losses in such accounts and does not believe it is exposed to significant risk on these funds.
Fair value measurement
The Company accounts for certain assets and liabilities at fair value and is required to disclose information that enables an assessment of the inputs used in determining the reported fair values. The fair value hierarchy prioritizes valuation inputs based on the observable nature of those inputs. The hierarchy applies only to the valuation inputs used to determine the reported fair value of the investments and is not a measure of the investment credit quality. The hierarchy defines three levels of valuation inputs:
Level 1 – Quoted prices in active markets for identical assets or liabilities.
Level 2 – Inputs other than quoted prices included within Level 1 that are observable for the asset or liability, either directly or indirectly.
Level 3 – Unobservable inputs that reflect the Company’s own assumptions about the assumptions market participants would use in pricing the asset or liability.
The Company’s financial instruments include cash and cash equivalents, short- and long-term marketable securities, accounts payable, contingent consideration, success payment liabilities, and other accrued liabilities. The carrying amounts of cash, cash equivalents, accounts payable, and accrued liabilities approximate fair value due to the short-term nature of these instruments. To the extent the valuation of financial instruments is based on models or inputs that are less observable or unobservable in the market, the determination of fair value requires more judgment. Accordingly, the degree of judgment exercised by the Company in determining fair value is greatest for instruments categorized in Level 3. See Note 6, Fair value measurements for more information on how the Company determines fair value.
Property and equipment, net
Property and equipment are stated at cost, net of accumulated depreciation. Depreciation of property and equipment is computed using the straight-line method over the estimated useful lives of the respective assets, generally
163
resulting gain or loss is recorded in other income (expense), net in the period realized. Maintenance and repairs are expensed as incurred.
Impairment of long-lived assets
The Company reviews the carrying value and estimated lives of its long-lived assets whenever events or circumstances indicate the carrying values may not be recoverable. Should an impairment exist, the impairment loss would be measured based on the excess of the asset’s carrying amount over its fair value. The Company has not recognized any impairment losses since inception.
Acquisitions
The Company accounts for business combinations using the acquisition method of accounting, which requires the assets acquired, including in-process research and development (IPR&D), and liabilities assumed be recorded at fair value as of the acquisition date. Any excess of the purchase price over the fair value of net assets acquired is recorded as goodwill. The determination of the estimated fair value of these items requires significant estimates and assumptions. Transaction costs associated with business combinations are recorded in general and administrative expense as they are incurred.
If the Company determines the acquisition does not meet the definition of a business combination under the acquisition method of accounting, the transaction is accounted for as an asset acquisition. In an asset acquisition, up-front payments allocated to IPR&D are recorded in research and development expense if it is determined that there is no alternative future use, and subsequent milestone payments are recorded in research and development expense when achieved.
Goodwill and intangible assets
Goodwill represents the excess of the purchase price over the estimated fair value of the identifiable assets acquired and liabilities assumed in a business combination. The Company evaluates goodwill for impairment annually or when a triggering event occurs that could indicate a potential impairment. The evaluation for impairment includes assessing qualitative factors or performing a quantitative analysis to determine whether it is more-likely-than-not that the fair value of net assets is below the carrying amount. As of December 31, 2021, the Company had goodwill of $
Intangible assets acquired in a business combination are recognized separately from goodwill and are initially recognized at fair value at the acquisition date. The fair value of the IPR&D is estimated using the replacement cost method. Under this method, the Company estimates the cost to recreate the technology and derive an estimated value to develop the technology. IPR&D assets are required to be classified as indefinite-lived assets and are not amortized until they become finite-lived assets upon the successful completion of the associated research and development technology. At that time, the useful life of the asset will be determined, and amortization will begin. If the associated research and development technology is abandoned, the related IPR&D asset will be written off and an impairment charge recorded. Intangible assets are reviewed for impairment at least annually or when a triggering event occurs that could indicate a potential impairment. As of December 31, 2021, there was
Contingent consideration from business combinations
Contingent consideration from a business combination is recorded at fair value on the acquisition date and remeasured at each subsequent reporting period with changes in fair value recognized in research and development related success payments and contingent consideration. Changes in fair values reflect changes to the Company’s assumptions regarding probabilities of successful achievement of related milestones, the timing in which the milestones are expected to be achieved, and the discount rate used to estimate the fair value of the obligation.
Success payments
The Company granted rights to a success payment to Cobalt Biomedicine, Inc. (Cobalt) pursuant to the terms of its acquisition agreement and to the President and Fellows of Harvard College (Harvard) pursuant to the terms of its exclusive license agreement. See Note 3, Acquisitions and Note 4, License and collaboration agreements for more details on the success payments.
The success payments are accounted for under Accounting Standards Codification (ASC) 815, Derivatives and Hedging. The success payment related to the acquisition of Cobalt (Cobalt Success Payment) was recorded as a liability on the consolidated balance sheet at fair value on the acquisition date and is remeasured at each subsequent reporting period, with changes in fair value recognized
164
in research and development related success payments and contingent consideration. For the success payments to Harvard (Harvard Success Payments), both the initial value and subsequent changes in fair value are recorded in research and development related success payments and contingent consideration.
To determine the estimated fair value of the success payment liabilities, the Company uses a Monte Carlo simulation methodology, which models the value of the liabilities based on several key assumptions, including the remaining terms of the success payments, risk-free interest rate, estimated number and timing of valuation measurement dates on the basis of which payments may be triggered, and expected volatility of the Company’s common stock. Expected volatility is estimated using the volatility of peer companies for a period of time commensurate with the remaining terms of the success payments. Additionally, prior to the IPO, the Cobalt Success Payment liability incorporated the estimated future value of the Company implied by the estimated per share value of the Company’s Series B convertible preferred stock at issuance, and the Harvard Success Payment liability incorporated the estimated per share value of the Company’s Series A convertible preferred stock. Concurrent with the closing of the Company’s IPO in February 2021, the Company’s Series A and Series B convertible preferred stock converted into common stock. Subsequent to the IPO, the computation of the estimated fair value of the Cobalt Success Payment liability incorporates the market capitalization of the Company at the end of each reporting period, and the computation of the estimated fair value of the Harvard Success Payments incorporates the per share fair market value of the Company’s common stock at the end of each reporting period.
Leases
At the inception of the arrangement, the Company determines whether the arrangement is or contains a lease based on the unique facts and circumstances present in the arrangement. Lease liabilities represent an obligation to make payments arising from a lease and are measured at the present value of the remaining future lease payments over the term of the lease. The present value of the lease payments is determined using an incremental borrowing rate (IBR), which reflects the fixed rate at which the Company could borrow the amount of the lease payments, on a collateralized basis, for a similar term and economic environment. The lease terms may include the impact of options to extend or terminate the lease when it is reasonably certain that the Company will exercise the option. Assumptions made by the Company at the commencement date are re-evaluated upon the occurrence of certain events, including a lease modification. When a lease modification results in a separate contract, it is accounted for in the same manner as a new lease. Right-of-use (ROU) assets represent the right to use the underlying asset identified in the lease for the term of the agreement. The calculation of the ROU asset incorporates the value of the lease liability and excludes any lease incentives received and initial direct costs incurred.
The Company’s lease portfolio consists of operating leases related to its facilities for office, laboratory, and manufacturing space. The Company does not have any financing leases. Leases with a term of 12 months or less are considered short-term and do not require recognition on the balance sheet, and payments associated with short-term leases are expensed as incurred. Rent expense for operating leases is recognized on a straight-line basis over the lease term.
Claims and contingencies
From time to time, the Company may become involved in litigation and proceedings relating to claims arising from the ordinary course of business. The Company accrues a liability if the likelihood of an adverse outcome is probable, and the amount can be reasonably estimated. If the likelihood of an adverse outcome is only reasonably possible, or if an adverse outcome is probable, but an estimate is not determinable, the Company provides disclosure of the material claim or contingency.
Convertible preferred stock
The carrying value of the Company’s Series A-1, Series A-2, and Series B convertible preferred stock was adjusted to reflect dividends if and when declared by the Company’s board of directors.
Stock-based compensation
The Company recognizes compensation costs related to restricted stock awards (RSAs), restricted stock units (RSUs), and stock options granted to employees and nonemployees based on the estimated fair value of the awards on the date of grant and recognizes expense on a straight-line basis over the requisite service period, which is generally the vesting period of the award. Forfeitures are recognized as they occur. For RSAs and RSUs, the fair value of the Company’s common stock is used to determine the resulting
165
stock-based compensation expense. The fair value of stock options is estimated on the date of grant using a Black-Scholes option pricing model which requires management to apply judgment and make estimates, including:
|
|
• |
Fair Value of Common Stock—The fair value of common stock is based on the closing price as reported on The Nasdaq Global Select Market on the date of grant. |
|
|
• |
Expected Term—The expected term represents the period that a stock-based award is expected to be outstanding. The Company uses the simplified method to determine the expected term, which is based on the average of the time-to-vesting and the contractual life of the option. |
|
|
• |
Expected Volatility— Due to the Company’s limited operating history and lack of company-specific historical and implied volatility data, the expected volatility is estimated based on the average historical volatilities of common stock of comparable publicly traded entities over a period of time commensurate with the expected term of the stock option grants. The comparable companies are chosen based on their size, stage in the product development cycle, or area of specialty. The Company will continue to apply this process until sufficient historical information regarding the volatility of its own stock price becomes available. |
|
|
• |
Risk-Free Interest Rate—The risk-free interest rate is based on the U.S. Treasury yield in effect at the time of grant for zero-coupon U.S. Treasury notes with maturities approximately equal to the expected term of the awards. |
|
|
• |
Expected Dividend—The Company has never paid dividends on its common stock and has no plans to pay dividends on its common stock. Therefore, the Company used an expected dividend yield of |
Research and development expense
The Company records expense for research and development costs as incurred. Nonrefundable, advance payments for goods or contracts for services are deferred, and expense is recognized in the period in which the goods are received or the services are rendered. Research and development expense consist of personnel-related costs, including salaries, benefits, and non-cash stock-based compensation, external research and development expenses incurred under arrangements with third parties, costs for laboratory supplies, costs to acquire and license technologies aligned with our goal of translating engineered cells to medicines, facility and other allocated expenses, including rent, depreciation, and allocated overhead costs, and other research and development expenses.
Research and development related success payment and contingent consideration
Research and development related success payments and contingent consideration include the change in the estimated fair value of the Cobalt Success Payment and Harvard Success Payment liabilities and Cobalt Contingent Consideration. Research and development expense related to the success payment liabilities and contingent consideration is unpredictable and may vary significantly from quarter-to-quarter and year-to-year due to changes in the assumptions used in the calculations.
General and administrative expenses
General and administrative expenses consist of personnel costs, including salaries, benefits, and non-cash stock-based compensation, for employees in finance, legal, executive, human resources, information technology, and other administrative functions, legal and consulting fees, recruiting costs, and facility costs not otherwise included in research and development expenses. Legal fees include those related to corporate and patent matters.
Income taxes
The Company determines its deferred tax assets and liabilities based on the differences between the financial statement and tax bases of assets and liabilities using enacted tax rates in effect for the year in which the differences are expected to reverse. A valuation allowance is recorded when it is more likely than not that the deferred tax asset will not be recovered. The Company applies judgment in the determination of the financial statement recognition and measurement of a tax position taken or expected to be taken in a tax return. The Company recognizes any material interest and penalties related to unrecognized tax benefits in income tax expense.
The Company is required to file income tax returns in the United States (U.S.) federal jurisdiction, and other state and local jurisdictions. The Company is generally subject to examination by U.S. federal and local income tax authorities for all tax years in which the loss carryforward is available. The Company is currently not under examination by the Internal Revenue Service or other jurisdictions for any tax years.
166
Segments
Operating segments are identified as components of an enterprise about which separate discrete financial information is available for evaluation by the chief operating decision-maker in making decisions regarding resource allocation and assessing performance. The Company views its operations and manages its business in
JOBS Act accounting election
The Company is an emerging growth company (EGC), as defined in the Jumpstart Our Business Startups Act of 2012 (JOBS Act). Under the JOBS Act, an EGC can delay adopting new or revised accounting standards issued subsequent to the enactment of the JOBS Act until such time as those standards apply to private companies. The Company has elected to use this extended transition period for complying with new or revised accounting standards that have different effective dates for public and private companies; however, the Company may adopt new or revised accounting standards early if the standard allows for early adoption.
In addition, the Company will utilize other exemptions and reduced reporting requirements provided to EGCs by the JOBS Act. Subject to certain conditions set forth in the JOBS Act, an EGC is not required to, among other things, (i) provide an auditor’s attestation report on the Company’s system of internal controls over financial reporting pursuant to Section 404(b) of the Sarbanes-Oxley Act of 2002, (ii) provide all of the compensation disclosure that may be required of non-emerging growth public companies under the Dodd-Frank Wall Street Reform and Consumer Protection Act, (iii) comply with any requirement that may be adopted by the Public Company Accounting Oversight Board regarding mandatory audit firm rotation or a supplement to the auditor’s report providing additional information about the audit and the financial statements (auditor discussion and analysis), or (iv) disclose certain executive compensation-related items, such as the correlation between executive compensation and performance and comparisons of the chief executive officer’s compensation to median employee compensation.
Recent accounting pronouncements
Recently adopted
In June 2016, the Financial Accounting Standards Board (FASB) issued Accounting Standards Update (ASU) 2016-13, Financial Instruments—Credit Losses (Topic 362): Measurement of Credit Losses on Financial Statements (ASU 2016-13). The new standard requires that expected credit losses relating to financial assets measured on an amortized cost basis and available-for-sale debt securities be recorded through an allowance for credit losses. It also limits the amount of credit losses to be recognized for available-for-sale debt securities to the amount by which the carrying value exceeds fair value and also requires the reversal of previously recognized credit losses if fair value increases. The targeted transition relief standard allows companies an option to irrevocably elect the fair value option of ASC 825-10, Financial Instruments-Overall, applied on an instrument-by-instrument basis for eligible instruments. The Company adopted ASU 2016-13 effective
Not yet adopted
ASU No. 2017-04, Intangibles—Goodwill and Other (Topic 350): Simplifying the Test for Goodwill Impairment
In January 2017, the FASB issued ASU 2017-04, Intangibles—Goodwill and Other (Topic 350): Simplifying the Test for Goodwill Impairment (ASU 2017-04). To address concerns over the cost and complexity of the two-step goodwill impairment test, the amendments in this ASU remove the second step of the test. An entity will instead apply a one-step quantitative test and record the amount of goodwill impairment as the excess of a reporting unit’s carrying amount over its fair value, not to exceed the total amount of goodwill allocated to the reporting unit. The new guidance does not amend the optional qualitative assessment of goodwill impairment. The new standard will be effective beginning January 1, 2023. The adoption of ASU 2017-04 is not expected to have a material impact on the Company’s consolidated financial statements.
3. Acquisitions
Oscine Corp.
In September 2020, the Company entered into a stock purchase agreement to acquire
167
The Company evaluated the acquisition and determined the screen test, as permitted under ASC 805, Business Combinations, was met, as the $
The Company is required to make up to an aggregate of $
Cytocardia, Inc.
In November 2019, the Company acquired
The Company evaluated the acquisition and determined the screen test, as permitted under ASC 805, Business Combinations, was met, as the $
The Company is required to make future milestone payments of up to an aggregate of $
Cobalt Biomedicine, Inc.
In February 2019, the Company acquired
168
The Company accounted for the Cobalt acquisition as a business combination using the acquisition method of accounting. Under this method, the assets acquired and liabilities assumed are measured at their fair values as of the acquisition date. The elements of the purchase consideration are as follows (in thousands):
|
Series A-2 convertible preferred stock issued(1) |
|
$ |
|
|
|
First milestone - restricted Series A-2 convertible preferred stock(2) |
|
|
|
|
|
Success payment(3) |
|
|
|
|
|
Fair value of contingent consideration(4) |
|
|
|
|
|
Other |
|
|
|
|
|
Total consideration |
|
$ |
|
|
|
|
(1) |
The purchase consideration included |
|
|
(2) |
The Company concluded the value of the first milestone, to be paid in restricted shares of Series A-2 convertible preferred stock, met the definition of being indexed to common stock. The restricted Series A-2 convertible preferred shares were recorded in convertible preferred stock valued at $ |
|
|
(3) |
The fair value of the success payment was determined using a Monte Carlo simulation methodology, which models the estimated fair value of the liability based on several key assumptions including the term of the success payment, expected volatility, risk-free interest rate, estimated number and timing of valuation measurement dates on the basis of which payment may be triggered, and the estimated future value of the Company implied by the estimated future per share value of the Company’s Series B convertible preferred stock at issuance. |
|
|
(4) |
The fair value of the contingent consideration was determined by calculating the probability-weighted value of the milestone payments based on the assessment of the likelihood and estimated timing that certain milestones would be achieved and using estimated discount rates ranging from |
The allocation of the purchase price is based on the estimated fair value of the assets acquired and liabilities assumed as of the date of acquisition. The components of the purchase price allocation are as follows (in thousands):
|
Net working capital |
|
$ |
( |
) |
|
Property and equipment |
|
|
|
|
|
Net liabilities assumed |
|
|
( |
) |
|
Deferred tax liability |
|
|
( |
) |
|
Acquired in-process research and development |
|
|
|
|
|
Goodwill |
|
|
|
|
|
Total consideration |
|
$ |
|
|
As part of the Cobalt acquisition, the Company recorded an intangible asset of $
The Company recognized $
Pursuant to the terms and conditions in the Cobalt acquisition agreement, the Company has an obligation to pay to certain former Cobalt stockholders up to an aggregate of $
In addition to our IPO, a valuation measurement date would be triggered upon a change of control of the Company if at least one Company product based on the fusogen technology is the subject of an active research program at the time of such change of control. If there is a change of control and the Company’s market capitalization is below $
169
the amount of the potential Cobalt Success Payment will decrease, and the amount of potential Cobalt Contingent Consideration will increase.
The following table sets forth various thresholds for the Company’s market capitalizations as of the date of a change of control and the resulting potential Cobalt Success Payment and additional potential Cobalt Contingent Consideration:
|
Sana market capitalization upon a change of control and resulting impact to Cobalt Success Payment and additional potential Cobalt Contingent Consideration |
|
Cobalt Success Payment |
|
|
Additional potential Cobalt Contingent Consideration |
|
||
|
|
|
(in millions) |
|
|||||
|
Equal to or exceeds $ |
|
$ |
|
|
|
$ |
- |
|
|
Equal to or exceeds $ |
|
|
|
|
|
|
|
|
|
Equal to or exceeds $ |
|
|
|
|
|
|
|
|
|
Less than $ |
|
|
- |
|
|
|
|
|
The Cobalt Success Payment and Cobalt Contingent Consideration liabilities are carried at fair value with changes in fair value recognized in research and development related success payments and contingent consideration. As of December 31, 2021 and 2020, the estimated fair value of the Cobalt Success Payment liability was $
4. License and collaboration agreements
Beam Therapeutics Inc.
In October 2021, the Company entered into an option and license agreement with Beam Therapeutics Inc. (Beam), pursuant to which the Company was granted a non-exclusive license to use Beam’s proprietary CRISPR Cas12b nuclease editing technology to research, develop, and commercialize engineered cell therapy products that (i) are directed to certain antigen targets, with respect to the Company’s allogeneic T cell programs, or (ii) comprise certain human cell types, with respect to the Company’s stem cell-derived programs. The Company made an upfront payment of $
President and Fellows of Harvard College
In March 2019, the Company entered into an exclusive license agreement with Harvard to access certain intellectual property for the development of hypoimmune cells. Under this agreement, the Company paid $
In connection with this agreement, the Company agreed to pay Harvard a license payment of $
Under the terms of the agreement, the Company may be required to pay up to an aggregate of $
170
the Company’s common stock relative to the original issuance price of $
The aggregate amount of the Harvard Success Payments will not exceed an aggregate of $
|
Multiple of Equity Value at Issuance |
|
5x |
|
|
10x |
|
|
20x |
|
|
30x |
|
|
40x |
|
|||||
|
Per share common stock price required for payment |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
Success payment(s) (in millions) |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
The Harvard Success Payment liabilities are carried at fair value, with the initial value and changes in fair value recognized in the consolidated statements of operations in research and development related success payments and contingent consideration. As of December 31, 2021 and 2020, the estimated fair value of the Harvard Success Payment liability was $
5. Restricted cash
As of December 31, 2021 and 2020, the Company maintained
|
|
|
December 31, |
|
|||||
|
|
|
2021 |
|
|
2020 |
|
||
|
|
|
(in thousands) |
|
|||||
|
Fremont, CA |
|
$ |
|
|
|
$ |
- |
|
|
Cambridge, MA |
|
|
|
|
|
|
|
|
|
South San Francisco, CA |
|
|
|
|
|
|
|
|
|
Seattle, WA |
|
|
|
|
|
|
|
|
|
Total restricted cash |
|
$ |
|
|
|
$ |
|
|
171
6. Fair value measurements
The following tables summarize the Company’s financial assets and liabilities measured at fair value on a recurring basis based on the three-tier fair value hierarchy:
|
|
|
|
|
December 31, 2021 |
|
|||||||||||||
|
|
|
Valuation Hierarchy |
|
Amortized Cost |
|
|
Gross Unrealized Holding Gains |
|
|
Gross Unrealized Holding Losses |
|
|
Estimated Fair Value |
|
||||
|
|
|
|
|
(in thousands) |
|
|||||||||||||
|
Financial assets: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cash equivalents: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Money market funds |
|
Level 1 |
|
$ |
|
|
|
$ |
- |
|
|
$ |
- |
|
|
$ |
|
|
|
Corporate debt securities |
|
Level 2 |
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Total cash equivalents |
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Short-term marketable securities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
U.S. government and agency securities |
|
Level 2 |
|
|
|
|
|
|
|
|
|
|
( |
) |
|
|
|
|
|
Corporate debt securities |
|
Level 2 |
|
|
|
|
|
|
- |
|
|
|
( |
) |
|
|
|
|
|
Total short-term marketable securities |
|
|
|
|
|
|
|
|
|
|
|
|
( |
) |
|
|
|
|
|
Long-term marketable securities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
U.S. government and agency securities |
|
Level 2 |
|
|
|
|
|
|
- |
|
|
|
( |
) |
|
|
|
|
|
Corporate debt securities |
|
Level 2 |
|
|
|
|
|
|
- |
|
|
|
( |
) |
|
|
|
|
|
Total long-term marketable securities |
|
|
|
|
|
|
|
|
- |
|
|
|
( |
) |
|
|
|
|
|
Other assets |
|
Level 3 |
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Total financial assets |
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
( |
) |
|
$ |
|
|
|
Financial liabilities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Short-term financial liabilities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Contingent consideration |
|
Level 3 |
|
$ |
|
|
|
$ |
- |
|
|
$ |
- |
|
|
$ |
|
|
|
Success payment liabilities |
|
Level 3 |
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Total short-term financial liabilities |
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Long-term financial liabilities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Contingent consideration |
|
Level 3 |
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Success payment liabilities |
|
Level 3 |
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Total long-term financial liabilities |
|
|
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Total financial liabilities |
|
|
|
$ |
|
|
|
$ |
- |
|
|
$ |
- |
|
|
$ |
|
|
172
|
|
|
|
|
December 31, 2020 |
|
|||||||||||||
|
|
|
Valuation Hierarchy |
|
Amortized Cost |
|
|
Gross Unrealized Holding Gains |
|
|
Gross Unrealized Holding Losses |
|
|
Estimated Fair Value |
|
||||
|
|
|
|
|
(in thousands) |
|
|||||||||||||
|
Financial assets: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cash equivalents: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Money market funds |
|
Level 1 |
|
$ |
|
|
|
$ |
- |
|
|
$ |
- |
|
|
$ |
|
|
|
U.S. government and agency securities |
|
Level 2 |
|
|
|
|
|
|
|
|
|
|
( |
) |
|
|
|
|
|
Corporate debt securities |
|
Level 2 |
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Total cash equivalents |
|
|
|
|
|
|
|
|
|
|
|
|
( |
) |
|
|
|
|
|
Short-term marketable securities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
U.S. government and agency securities |
|
Level 2 |
|
|
|
|
|
|
|
|
|
|
( |
) |
|
|
|
|
|
Corporate debt securities |
|
Level 2 |
|
|
|
|
|
|
- |
|
|
|
( |
) |
|
|
|
|
|
Total short-term marketable securities |
|
|
|
|
|
|
|
|
|
|
|
|
( |
) |
|
|
|
|
|
Long-term marketable securities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
U.S. government and agency securities |
|
Level 2 |
|
|
|
|
|
|
|
|
|
|
- |
|
|
|
|
|
|
Total long-term marketable securities |
|
|
|
|
|
|
|
|
|
|
|
|
- |
|
|
|
|
|
|
Total financial assets |
|
|
|
$ |
|
|
|
$ |
|
|
|
$ |
( |
) |
|
$ |
|
|
|
Financial liabilities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Long-term financial liabilities: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Contingent consideration |
|
Level 3 |
|
$ |
|
|
|
$ |
- |
|
|
$ |
- |
|
|
$ |
|
|
|
Success payment liabilities |
|
Level 3 |
|
|
|
|
|
|
- |
|
|
|
- |
|
|
|
|
|
|
Total financial liabilities |
|
|
|
$ |
|
|
|
$ |
- |
|
|
$ |
- |
|
|
$ |
|
|
The Company measures the fair value of money market funds based on quoted prices in active markets for identical assets or liabilities. The Level 2 marketable securities include U.S. government, agency securities, and corporate debt securities and are valued based on either recent trades of securities in inactive markets or quoted market prices of similar instruments and other significant inputs derived from or corroborated by observable market data. As of December 31, 2021, all marketable securities had an effective maturity date of
Securities in an unrealized loss position have been in an unrealized loss position for less than one year. The Company determined that there was no material change in the credit risk of the above investments during the year ended December 31, 2021. As such, an allowance for credit losses has not been recognized. As of December 31, 2021, the Company does not intend to sell such securities, and it is not more-likely-than-not that the Company will be required to sell the securities prior to the recovery of the amortized cost basis.
As of December 31, 2021 and 2020, the balance in accumulated other comprehensive income (loss) included net unrealized gains (losses) related to the Company’s available-for-sale debt securities. There were
The following table sets forth a summary of the changes in the fair value of the Company’s Level 3 financial liabilities:
|
|
|
Contingent Consideration |
|
|
Cobalt Success Payment Liability |
|
|
Harvard Success Payment Liability |
|
|||
|
|
|
(in thousands) |
|
|||||||||
|
Balance as of December 31, 2020 |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
Changes in fair value - expense (gain) |
|
|
|
|
|
|
|
|
|
|
|
|
|
Balance as of March 31, 2021 |
|
|
|
|
|
|
|
|
|
|
|
|
|
Changes in fair value - expense (gain) |
|
|
|
|
|
|
( |
) |
|
|
( |
) |
|
Balance as of June 30, 2021 |
|
|
|
|
|
|
|
|
|
|
|
|
|
Changes in fair value - expense (gain) |
|
|
( |
) |
|
|
|
|
|
|
|
|
|
Balance as of September 30, 2021 |
|
|
|
|
|
|
|
|
|
|
|
|
|
Changes in fair value - expense (gain) |
|
|
|
|
|
|
( |
) |
|
|
( |
) |
|
Balance as of December 31, 2021 |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
173
Contingent consideration
The Company utilizes significant estimates and assumptions it believes would be made by a market participant in determining the estimated fair value of the Cobalt Contingent Consideration at each balance sheet date. The fair value of the Cobalt Contingent Consideration was determined by calculating the probability-weighted estimated value of the pre-specified development milestone payments based on the assessment of the likelihood and estimated timing that the milestones would be achieved and the applicable discount rates. The discount rate captures the credit risk associated with the payment of the contingent consideration when earned and due. The Company assesses these estimates on an on-going basis as additional data impacting the assumptions are obtained.
The fair value of the Cobalt Contingent Consideration was calculated using the following unobservable inputs:
|
|
|
December 31, |
|
|||||||
|
|
|
2021 |
|
|
2020 |
|
||||
|
Unobservable Input |
|
Range |
|
Weighted-Average |
|
|
Range |
|
Weighted-Average |
|
|
Discount rates |
|
|
|
|
|
|
|
|
|
|
|
Probability of milestone achievement |
|
|
|
|
|
|
|
|
|
|
The weighted-average unobservable inputs were calculated based on the relative value of the pre-specified development milestones. The estimated fair value of the Cobalt Contingent Consideration may change significantly as development progresses and additional data are obtained, impacting the assumptions regarding probabilities of successful achievement of the milestones used to estimate the fair value of the liability and the timing in which they are expected to be achieved. In evaluating the fair value assumptions, judgment is required to interpret the market data used to develop the estimates. The estimates of fair value may not be indicative of the amounts that could be realized in a current market exchange. Accordingly, the use of different market assumptions, inputs, and/or different valuation techniques could result in materially different fair value estimates.
Success payments
The Company utilizes significant estimates and assumptions in determining the estimated fair value of the success payment liabilities and the associated expense or gain at each balance sheet date. The estimated fair value of the Cobalt and Harvard success payment liabilities was determined using a Monte Carlo simulation methodology, which models the estimated fair value of the liability based on several key assumptions, including the expected volatility, remaining term, risk-free interest rate, estimated number and timing of valuation measurement dates on the basis of which payment may be triggered, and for the Cobalt Success Payment, the Company’s market capitalization, and for the Harvard Success Payments, the per share fair value of the Company’s common stock.
Prior to the IPO, the calculation of the estimated fair value of the success payment liabilities incorporated the estimated future per share value of the Company’s Series A convertible preferred stock and the estimated future value of the Company implied by the estimated future per share value of the Company’s Series B convertible preferred stock at issuance. Concurrent with the closing of the Company’s IPO in February 2021, the Company’s Series A and Series B convertible preferred stock converted into common stock. As such, subsequent to the IPO, the computation of the estimated fair value of the Harvard Success Payment liabilities incorporates the per share fair market value of the Company’s common stock, and the estimated fair value of the Cobalt Success Payment liability incorporates the market capitalization of the Company.
The fair values of the Cobalt and Harvard success payments were calculated using the following unobservable inputs:
|
|
|
December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
||||||
|
Unobservable Input |
|
Cobalt |
|
|
Harvard |
|
|
Cobalt |
|
|
Harvard |
|
|
Expected stock price volatility |
|
|
|
|
|
|
|
|
|
|
|
|
|
Expected term (years) |
|
|
|
|
|
|
|
|
|
|
|
|
174
7. Property and equipment, net
Property and equipment, net consists of the following:
|
|
|
December 31, |
|
|||||
|
|
|
2021 |
|
|
2020 |
|
||
|
|
|
(in thousands) |
|
|||||
|
Laboratory equipment |
|
$ |
|
|
|
$ |
|
|
|
Leasehold improvements |
|
|
|
|
|
|
|
|
|
Construction in progress |
|
|
|
|
|
|
|
|
|
Computer equipment, software, and other |
|
|
|
|
|
|
|
|
|
Total property and equipment, at cost |
|
|
|
|
|
|
|
|
|
Less: Accumulated depreciation |
|
|
( |
) |
|
|
( |
) |
|
Property and equipment, net |
|
$ |
|
|
|
$ |
|
|
Depreciation expense was $
8. Accrued liabilities
Accrued compensation and accrued expenses and other current liabilities consist of the following:
|
|
|
December 31, |
|
|||||
|
|
|
2021 |
|
|
2020 |
|
||
|
|
|
(in thousands) |
|
|||||
|
Accrued compensation: |
|
|
|
|
|
|
|
|
|
Accrued bonus |
|
$ |
|
|
|
$ |
|
|
|
Accrued paid time off |
|
|
|
|
|
|
|
|
|
Accrued payroll |
|
|
|
|
|
|
|
|
|
Total accrued compensation |
|
$ |
|
|
|
$ |
|
|
|
Accrued expenses and other current liabilities: |
|
|
|
|
|
|
|
|
|
Accrued research and development expense services |
|
$ |
|
|
|
$ |
|
|
|
Accrued property and equipment |
|
|
|
|
|
|
|
|
|
Accrued professional fees |
|
|
|
|
|
|
|
|
|
Other accrued current liabilities |
|
|
|
|
|
|
|
|
|
Total accrued expenses and other current liabilities |
|
$ |
|
|
|
$ |
|
|
9. Commitments and contingencies
Lease commitments
The Company’s lease portfolio is primarily composed of operating leases for office, laboratory, non-good manufacturing practices (GMP) pilot plant manufacturing, and industrial space located in Seattle, WA, Cambridge, MA, South San Francisco, CA, and Fremont, CA. Our operating leases have contractual periods expiring between
175
capabilities, and include a lease incentive allowance.
|
Location |
|
Approximate Square Footage |
|
Commencement Dates |
|
Expiration Dates |
|
Seattle, WA |
|
|
|
|
|
|
|
Cambridge, MA |
|
|
|
|
|
|
|
South San Francisco, CA |
|
|
|
|
|
|
|
Fremont, CA |
|
|
|
|
|
November 2031 |
Throughout the term of the lease agreements, the Company is responsible for paying certain operating costs in addition to rent, such as common area maintenance, taxes, utilities, and insurance. These additional charges are considered variable lease costs and are recognized in the period in which the costs are incurred.
The following table summarizes the Company’s lease costs:
|
|
|
Year Ended December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
|
2019 |
|
|||
|
|
|
(in thousands) |
|
|||||||||
|
Operating lease cost |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
Short-term lease cost |
|
|
|
|
|
|
|
|
|
|
|
|
|
Variable lease cost |
|
|
|
|
|
|
|
|
|
|
|
|
|
Total lease cost |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
As of December 31, 2021, the weighted-average remaining lease term was
The following table reconciles the Company’s undiscounted operating lease cash flows by fiscal year to the present value of the operating lease liabilities as of December 31, 2021 (in thousands):
|
2022 |
|
|
|
|
|
2023 |
|
|
|
|
|
2024 |
|
|
|
|
|
2025 |
|
|
|
|
|
2026 |
|
|
|
|
|
2026 and thereafter |
|
|
|
|
|
Total undiscounted lease payments |
|
|
|
|
|
Less: imputed interest |
|
|
( |
) |
|
Less: tenant improvement allowances |
|
|
( |
) |
|
Present value of operating lease liabilities |
|
$ |
|
|
10. Convertible preferred stock
Series A-1, A-2, and B convertible preferred stock financings
In 2018 and 2019, the Company issued
In December 2020, the Company amended and restated its certificate of incorporation and amended the investors’ rights agreement and voting agreement with its stockholders. Under the Company’s amended and restated certificate of incorporation, the authorized capital stock of the Company increased to
11. Stockholders’ equity
Effective February 2021, the Company amended and restated its certificate of incorporation, increasing the number of shares of all classes of stock the Company has authority to issue to
176
As of December 31, 2021, there were
12. Stock-based compensation
2021 Incentive Award Plan
In February 2021, the Company adopted the 2021 Incentive Award Plan (2021 Plan), which became effective on the completion of the Company’s IPO. The 2021 Plan provides for a variety of stock-based compensation awards, including stock options, restricted stock awards (RSAs), and restricted stock units (RSUs). In conjunction with adopting the 2021 Plan, the Company discontinued the 2018 Equity Incentive Plan with respect to new equity awards.
The 2021 Plan provides for an annual increase in the shares available for issuance thereunder, to be added on the first day of each year, beginning in 2022 and ending in 2031, equal to the lesser of (i)
2021 Employee Stock Purchase Plan
In February 2021, the Company adopted the 2021 Employee Stock Purchase Plan (2021 ESPP), which became effective on the completion of the Company’s IPO. The 2021 ESPP allows eligible employees to purchase shares of the Company’s common stock at a discount through payroll deductions of up to
The 2021 ESPP also provides for an annual share increase, to be added on the first day of each year, beginning in 2022 and ending in 2031, equal to the lesser of (i)
Stock-based compensation expense
Stock-based compensation expense is recognized in the consolidated statements of operations as follows:
|
|
|
Year Ended December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
|
2019 |
|
|||
|
|
|
(in thousands) |
|
|||||||||
|
Research and development |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
General and administrative |
|
|
|
|
|
|
|
|
|
|
|
|
|
Total stock-based compensation expense |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
Unrecognized stock-based compensation costs related to unvested awards and the weighted-average period over which the costs are expected to be recognized as of December 31, 2021 are as follows:
|
|
|
Stock Options |
|
|
RSAs |
|
|
RSUs |
|
|||
|
Unrecognized stock-based compensation expense (in thousands) |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
Weighted-average period costs expected to be recognized (in years) |
|
|
|
|
|
|
|
|
|
|
|
|
177
Stock options
A summary of the Company’s stock option activity is as follows:
|
|
|
Stock Options (in thousands) |
|
|
Weighted-Average Exercise Price per Share |
|
|
Weighted-Average Remaining Contractual Life (in years) |
|
|
Aggregate Intrinsic Value (in thousands) |
|
||||
|
Outstanding as of December 31, 2020 |
|
|
|
|
|
$ |
|
|
|
|
|
|
|
|
|
|
|
Granted |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Exercised |
|
|
( |
) |
|
|
|
|
|
|
|
|
|
|
|
|
|
Forfeited/Cancelled |
|
|
( |
) |
|
|
|
|
|
|
|
|
|
|
|
|
|
Outstanding as of December 31, 2021 |
|
|
|
|
|
$ |
|
|
|
|
|
|
|
$ |
|
|
|
Exercisable as of December 31, 2021 |
|
|
|
|
|
$ |
|
|
|
|
|
|
|
$ |
|
|
The fair value of stock options granted to employees, directors, and consultants was estimated on the date of grant using the Black-Scholes option pricing model using the following assumptions:
|
|
|
Year Ended December 31, |
|
||||||
|
Assumptions |
|
2021 |
|
|
2020 |
|
|
2019 |
|
|
Risk free interest rate |
|
0.46% - 1.38% |
|
|
0.36% - 1.51% |
|
|
1.53% - 2.62% |
|
|
Expected volatility |
|
|
|
|
|
|
|
|
|
|
Expected term (years) |
|
5.50 - 6.40 |
|
|
6.25 - 6.75 |
|
|
6.02 - 6.25 |
|
|
Expected dividend |
|
|
|
|
|
|
|
|
|
The following table summarizes additional information related to stock option activity:
|
|
|
Year Ended December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
|
2019 |
|
|||
|
Weighted average grant date fair value per share for options granted |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
|
Aggregate intrinsic value of stock options exercised (in thousands) |
|
$ |
|
|
|
$ |
|
|
|
$ |
|
|
Restricted stock
A summary of the Company’s RSA and RSU activity is as follows:
|
|
|
RSAs (in thousands) |
|
|
RSAs Weighted-Average Grant Date Fair Value per Share |
|
|
RSUs (in thousands) |
|
|
RSUs Weighted-Average Grant Date Fair Value per Share |
|
||||
|
Unvested shares as of December 31, 2020 |
|
|
|
|
|
$ |
|
|
|
|
|
|
|
$ |
|
|
|
Granted |
|
|
- |
|
|
|
- |
|
|
|
|
|
|
|
|
|
|
Vested |
|
|
( |
) |
|
|
|
|
|
|
( |
) |
|
|
|
|
|
Forfeited |
|
|
( |
) |
|
|
|
|
|
|
( |
) |
|
|
|
|
|
Unvested shares as of December 31, 2021 |
|
|
|
|
|
$ |
|
|
|
|
|
|
|
$ |
|
|
The fair value of vested RSAs was $
13. Income taxes
As of December 31, 2021, the Company had U.S. federal and state tax-effected net operating loss (NOL) carryforwards of $
178
In connection with the 2019 Cobalt acquisition, the Company recorded a deferred tax liability of $
A reconciliation of income taxes computed using the U.S. federal statutory rate to that reflected in operations follows:
|
|
|
Year Ended December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
|
2019 |
|
|||
|
Federal statutory tax |
|
|
|
% |
|
|
|
% |
|
|
|
% |
|
State income tax, net of federal benefit |
|
|
|
|
|
|
|
|
|
|
|
|
|
Valuation allowance |
|
|
( |
) |
|
|
( |
) |
|
|
( |
) |
|
Success payment liabilities |
|
|
( |
) |
|
|
( |
) |
|
|
- |
|
|
Contingent consideration |
|
|
( |
) |
|
|
( |
) |
|
|
( |
) |
|
Tax credits |
|
|
|
|
|
|
|
|
|
|
|
|
|
Other |
|
|
|
|
|
|
( |
) |
|
|
( |
) |
|
Effective income tax rate |
|
|
|
% |
|
|
|
% |
|
|
|
% |
The principal components of the Company’s net deferred tax assets are as follows:
|
|
|
December 31, |
|
|||||
|
|
|
2021 |
|
|
2020 |
|
||
|
|
|
(in thousands) |
|
|||||
|
Deferred tax assets: |
|
|
|
|
|
|
|
|
|
Net operating loss carryforwards |
|
$ |
|
|
|
$ |
|
|
|
Lease liabilities |
|
|
|
|
|
|
|
|
|
Tax credit carryforwards |
|
|
|
|
|
|
|
|
|
Intangibles |
|
|
|
|
|
|
- |
|
|
Accrued liabilities and allowances |
|
|
|
|
|
|
|
|
|
Success payment liabilities |
|
|
|
|
|
|
|
|
|
Stock-based compensation |
|
|
|
|
|
|
- |
|
|
Other |
|
|
- |
|
|
|
|
|
|
Gross deferred tax assets |
|
|
|
|
|
|
|
|
|
Valuation allowance |
|
|
( |
) |
|
|
( |
) |
|
Deferred tax assets, net of valuation allowance |
|
|
|
|
|
|
|
|
|
Deferred tax liabilities: |
|
|
|
|
|
|
|
|
|
Right-of-use assets |
|
|
( |
) |
|
|
( |
) |
|
Fixed assets |
|
|
( |
) |
|
|
( |
) |
|
Intangibles |
|
|
- |
|
|
|
( |
) |
|
Stock-based compensation |
|
|
- |
|
|
|
( |
) |
|
Other |
|
|
( |
) |
|
|
- |
|
|
Deferred tax liabilities |
|
|
( |
) |
|
|
( |
) |
|
Net deferred taxes assets |
|
$ |
- |
|
|
$ |
- |
|
The valuation allowance relates primarily to net U.S. deferred tax assets from operating losses, research tax credit carryforwards, and amounts paid and accrued to enter into various agreements for which the tax treatment requires capitalization and amortization.
The Company maintains a full valuation allowance on its net U.S. deferred tax assets. The assessment regarding whether a valuation allowance is required considers both positive and negative evidence when determining whether it is more likely than not that deferred tax assets are recoverable. In making this assessment, significant weight is given to evidence that can be objectively verified. In its evaluation, the Company considered its cumulative loss in the first year of operation and its forecasted losses in the near term as significant negative evidence. Based upon a review of the four sources of income identified within ASC 740, Accounting for Income Taxes, the Company determined that the negative evidence outweighed the positive evidence, and a full valuation allowance on its net deferred tax assets should be maintained. The Company will continue to assess the realizability of its deferred tax assets going forward and will adjust the valuation allowance as needed.
The Company determines its uncertain tax positions based on a determination of whether and how much of the tax benefit the Company takes in its tax filings or positions is more likely than not to be sustained upon examination by the relevant income tax
179
authorities. The Company is generally subject to examination by U.S. federal and local income tax authorities for all tax years in which the loss carryforward is available. The Company applies judgment in its determination of the financial statement recognition and measurement of tax positions taken or expected to be taken in a tax return. As of December 31, 2021 and 2020, the Company’s uncertain tax positions were immaterial.
14. Net loss per share
Basic and diluted net loss per common share are calculated by dividing net loss by the weighted-average number of common shares outstanding during the period, without consideration for common stock equivalents. The Company was in a loss position for all periods presented, and basic net loss per share and diluted net loss per share are therefore the same for all periods, as the inclusion of all potential common securities outstanding would have been anti-dilutive.
The following table summarizes the calculation of basic and diluted net loss per share of common stock:
|
|
|
Year Ended December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
|
2019 |
|
|||
|
|
|
(in thousands, except per share amounts) |
|
|||||||||
|
Net loss |
|
$ |
( |
) |
|
$ |
( |
) |
|
$ |
( |
) |
|
Weighted-average number of common shares - basic and diluted |
|
|
|
|
|
|
|
|
|
|
|
|
|
Net loss per common share - basic and diluted |
|
$ |
( |
) |
|
$ |
( |
) |
|
$ |
( |
) |
The following securities were excluded from the computation of net loss per diluted share of common stock for periods presented as their effect would have been anti-dilutive:
|
|
|
Year Ended December 31, |
|
|||||||||
|
|
|
2021 |
|
|
2020 |
|
|
2019 |
|
|||
|
|
|
(in thousands) |
|
|||||||||
|
Convertible preferred stock |
|
|
- |
|
|
|
|
|
|
|
|
|
|
Options to purchase common stock |
|
|
|
|
|
|
|
|
|
|
|
|
|
Unvested restricted common stock |
|
|
|
|
|
|
|
|
|
|
|
|
|
Unvested RSUs |
|
|
|
|
|
|
|
|
|
|
|
|
|
Total |
|
|
|
|
|
|
|
|
|
|
|
|
15. Employee benefit plan
In January 2019, the Company adopted a 401(k) retirement and savings plan (the 401(k) Plan) covering all employees. The 401(k) Plan allows employees to make pre- and post-tax contributions up to the maximum allowable amount set by the IRS. The Company has
180
Item 9. Changes in and Disagreements with Accountants on Accounting and Financial Disclosure.
None.
Item 9A. Controls and Procedures.
Evaluation of Disclosure Controls and Procedures
Our management, with the participation and supervision of our Chief Executive Officer and Chief Financial Officer, have evaluated the effectiveness of our disclosure controls and procedures as defined in Rules 13a-15(e) and 15d-15(e) of the Exchange Act, as of the end of the period covered by this Annual Report. Based on this evaluation, our Chief Executive Officer and Chief Financial Officer concluded that as of such date, our disclosure controls and procedures were effective at a reasonable assurance level.
Management’s Annual Report on Internal Control Over Financial Reporting
Management is responsible for establishing and maintaining adequate internal control over financial reporting, as defined in Rule 13a-15(f) under the Exchange Act. Our internal control over financial reporting is designed to provide reasonable assurance regarding the reliability of financial reporting and the preparation of consolidated financial statements for external purposes in accordance with GAAP. Because of its inherent limitations, our internal control over financial reporting may not prevent or detect misstatements. Also, projections of any evaluation of effectiveness to future periods are subject to the risk that controls may become inadequate because of changes in conditions, or that the degree of compliance with applicable policies or procedures may deteriorate.
Management has assessed the effectiveness of our internal control over financial reporting based on the framework set forth by the Committee of Sponsoring Organizations of the Treadway Commission in Internal Control-Integrated Framework. Based on our evaluation, management has concluded that our internal control over financial reporting was effective as of December 31, 2021. Management reviewed the results of this assessment with our audit committee.
Attestation Report of the Registered Public Accounting Firm
This Annual Report does not include an attestation report of our registered public accounting firm due to an exemption established by the JOBS Act for “emerging growth companies.”
Changes in Internal Control Over Financial Reporting
There were no changes in our internal control over financial reporting that occurred during the three months ended December 31, 2021 that materially affected, or are reasonably likely to materially affect, our internal control over financial reporting.
Item 9B. Other Information.
None.
Item 9C. Disclosure Regarding Foreign Jurisdictions that Prevent Inspections.
Not applicable.
181
PART III
Item 10. Directors, Executive Officers and Corporate Governance.
The information required by this item is incorporated by reference to the information in our Proxy Statement in the sections titled “Information About Our Board of Directors,” “Information About Our Executive Officers Who Are Not Directors,” and “Corporate Governance.”
Item 11. Executive Compensation.
The information required by this item is incorporated by reference to the information in our Proxy Statement in the sections titled “Executive Compensation,” “Director Compensation,” and “Corporate Governance – Committees of the Board of Directors – Compensation Committee Interlocks and Insider Participation.”
Item 12. Security Ownership of Certain Beneficial Owners and Management and Related Stockholder Matters.
The information required by this item is incorporated by reference to the information in our Proxy Statement in the sections titled “Securities Authorized For Issuance Under Equity Compensation Plans” and “Security Ownership of Certain Beneficial Owners and Management.”
Item 13. Certain Relationships and Related Transactions, and Director Independence.
The information required by this item is incorporated by reference to the information in our Proxy Statement in the sections titled “Corporate Governance – Board of Directors Independence” and “Transactions With Related Persons.”
Item 14. Principal Accountant Fees and Services.
The information required by this item is incorporated by reference to the information in our Proxy Statement in the sections titled “Independent Registered Public Accounting Firm Fees and Services” and “Pre-Approval Policies and Procedures.”
182
PART IV
Item 15. Exhibits, Financial Statement Schedules.
Exhibit Index
|
|
|
|
|
|
|
Incorporated by Reference |
|
|
||
|
Exhibit Number |
|
Exhibit Description |
|
Form |
|
Date |
|
Number |
|
Filed Herewith |
|
|
|
|
|
|
|
|
|
|
|
|
|
3.1 |
|
|
8-K |
|
02/08/2021 |
|
3.1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3.2 |
|
|
8-K |
|
02/08/2021 |
|
3.2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4.1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4.2 |
|
|
S-1/A |
|
01/28/2021 |
|
4.2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4.3 |
|
|
10-K |
|
03/24/2021 |
|
4.3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.1 |
|
|
S-1 |
|
01/13/2021 |
|
10.1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.2 |
|
Form of Indemnification Agreement for directors and officers |
|
S-1/A |
|
01/28/2021 |
|
10.2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.3(a)# |
|
|
10-Q |
|
11/8/2021 |
|
10.3(a) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.3(b)# |
|
Second Amendment to 2018 Equity Incentive Plan, dated as of November 9, 2020 |
|
S-1 |
|
01/13/2021 |
|
10.3(b) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.3(c)# |
|
Third Amendment to 2018 Equity Incentive Plan, dated as of December 4, 2020 |
|
S-1 |
|
01/13/2021 |
|
10.3(c) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.3(d)# |
|
Fourth Amendment to 2018 Equity Incentive Plan, dated as of December 8, 2021 |
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
10.3(e)# |
|
Form of Stock Option Agreement under 2018 Equity Incentive Plan |
|
S-1 |
|
01/13/2021 |
|
10.3(d) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.4(a)# |
|
|
S-1/A |
|
01/28/2021 |
|
10.4(a) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.4(b)# |
|
First Amendment to 2021 Incentive Award Plan, dated as of December 8, 2021 |
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
10.4(c)# |
|
Form of Stock Option Grant Notice and Stock Option Agreement under the 2021 Incentive Award Plan |
|
S-1/A |
|
01/28/2021 |
|
10.4(b) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.4(d)# |
|
Form of Stock Option Grant Notice and Stock Option Agreement under the 2021 Incentive Award Plan |
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
10.4(e)# |
|
|
S-1/A |
|
01/28/2021 |
|
10.4(c) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.4(f)# |
|
|
S-1/A |
|
01/28/2021 |
|
10.4(d) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.5# |
|
|
S-1/A |
|
01/28/2021 |
|
10.5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.6# |
|
|
S-1 |
|
01/13/2021 |
|
10.6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.7# |
|
|
S-1 |
|
01/13/2021 |
|
10.7 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.8# |
|
|
S-1 |
|
01/13/2021 |
|
10.8 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
183
|
|
|
|
|
|
|
Incorporated by Reference |
|
|
||
|
Exhibit Number |
|
Exhibit Description |
|
Form |
|
Date |
|
Number |
|
Filed Herewith |
|
10.9# |
|
|
S-1 |
|
01/13/2021 |
|
10.9 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.10# |
|
|
S-1 |
|
01/13/2021 |
|
10.10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.11# |
|
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.12(a) † |
|
|
S-1 |
|
01/13/2021 |
|
10.12(a) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.12(b) † |
|
|
S-1 |
|
01/13/2021 |
|
10.12(b) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.13(a) † |
|
|
S-1 |
|
01/13/2021 |
|
10.13(a) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.13(b) † |
|
|
S-1 |
|
01/13/2021 |
|
10.13(b) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.14† |
|
|
S-1 |
|
01/13/2021 |
|
10.14 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.15(a) † |
|
|
S-1 |
|
01/13/2021 |
|
10.15(a) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.15(b) † |
|
Amendment to License Agreement by and between the Company and Harvard, dated as of June 10, 2019 |
|
S-1 |
|
01/13/2021 |
|
10.15(b) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.15(c) † |
|
|
S-1 |
|
01/13/2021 |
|
10.15(c) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.15(d) † |
|
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.15(e) † |
|
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.16(a) † |
|
|
S-1 |
|
01/13/2021 |
|
10.16(a) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.16(b) † |
|
|
S-1 |
|
01/13/2021 |
|
10.16(b) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.17† |
|
|
S-1 |
|
01/13/2021 |
|
10.17 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.18† |
|
|
S-1 |
|
01/13/2021 |
|
10.18 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.19† |
|
|
S-1 |
|
01/13/2021 |
|
10.19 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.20(a) † |
|
|
S-1 |
|
01/13/2021 |
|
10.20(a) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
184
|
|
|
|
|
|
|
Incorporated by Reference |
|
|
||
|
Exhibit Number |
|
Exhibit Description |
|
Form |
|
Date |
|
Number |
|
Filed Herewith |
|
10.20(b) † |
|
|
S-1 |
|
01/13/2021 |
|
10.20(b) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.20(c) † |
|
|
S-1 |
|
01/13/2021 |
|
10.20(c) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.21(a) † |
|
|
S-1 |
|
01/13/2021 |
|
10.21(a) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.21(b) † |
|
|
S-1 |
|
01/13/2021 |
|
10.21(b) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.21(c) † |
|
|
S-1/A |
|
01/28/2021 |
|
10.21(c) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.21(d) † |
|
|
S-1 |
|
01/13/2021 |
|
10.21(c) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.21(e) † |
|
|
S-1/A |
|
01/28/2021 |
|
10.21(e) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.22# |
|
|
S-1 |
|
01/13/2021 |
|
10.22 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.23† |
|
|
S-1/A |
|
02/02/2021 |
|
10.23 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.24† |
|
|
10-Q |
|
11/08/2021 |
|
10.1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.25† |
|
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10.26† |
|
Lease Agreement by and between the Company and Pacific Commons Owner, LP, dated as of July 13, 2021 |
|
10-Q |
|
08/04/2021 |
|
10.1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
21.1 |
|
|
10-K |
|
03/24/2021 |
|
21.1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
23.1 |
|
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
24.1 |
|
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
31.1* |
|
Certificate of Principal Executive Officer Pursuant to Section 302 of the Sarbanes-Oxley Act of 2002 |
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
31.2* |
|
Certificate of Principal Financial Officer Pursuant to Section 302 of the Sarbanes-Oxley Act of 2002 |
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
32.1* |
|
Certificate of Principal Executive Officer Pursuant to Section 906 of the Sarbanes-Oxley Act of 2002 |
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
32.2* |
|
Certificate of Principal Financial Officer Pursuant to Section 906 of the Sarbanes-Oxley Act of 2002 |
|
|
|
|
|
|
|
X |
|
|
|
|
|
|
|
|
|
|
|
|
|
101.INS |
|
Inline XBRL Instance Document |
|
|
|
|
|
|
|
|
|
101.SCH |
|
Inline XBRL Taxonomy Extension Schema Document |
|
|
|
|
|
|
|
|
|
101.CAL |
|
Inline XBRL Taxonomy Extension Calculation Linkbase Document |
|
|
|
|
|
|
|
|
|
101.DEF |
|
Inline XBRL Taxonomy Extension Definition Linkbase Document |
|
|
|
|
|
|
|
|
|
101.LAB |
|
Inline XBRL Taxonomy Extension Label Linkbase Document |
|
|
|
|
|
|
|
|
185
|
|
|
|
|
|
|
Incorporated by Reference |
|
|
||
|
Exhibit Number |
|
Exhibit Description |
|
Form |
|
Date |
|
Number |
|
Filed Herewith |
|
101.PRE |
|
Inline XBRL Taxonomy Extension Presentation Linkbase Document |
|
|
|
|
|
|
|
|
|
104 |
|
Cover Page Interactive Data File (embedded within the Inline XBRL document) |
|
|
|
|
|
|
|
|
|
# |
Indicates management contract or compensatory plan. |
|
† |
Certain portions of this document that constitute confidential information have been redacted in accordance with Regulation S-K, Item 601(b)(10). |
|
* |
These certifications are not deemed filed with the Securities and Exchange Commission and are not to be incorporated by reference into any filing of the Company under the Securities Act of 1933, as amended, or the Securities Exchange Act of 1934, as amended, whether made before or after the date of this Annual Report, irrespective of any general incorporation language contained in such filing. |
Item 16. Form 10-K Summary
Not applicable.
186
SIGNATURES
Pursuant to the requirements of Section 13 or 15(d) of the Securities Exchange Act of 1934, as amended, the Registrant has duly caused this Annual Report on Form 10-K to be signed on its behalf by the undersigned, thereunto duly authorized in the City of Seattle, State of Washington on the 16th day of March 2022.
|
|
SANA BIOTECHNOLOGY, INC. |
|
|
|
|
|
|
|
/s/ Steven D. Harr, M.D. |
|
|
|
Name: |
Steven D. Harr, M.D. |
|
|
Title: |
President and Chief Executive Officer (Principal Executive Officer) |
|
|
|
|
|
|
/s/ Nathan Hardy |
|
|
|
Name: |
Nathan Hardy |
|
|
Title: |
Chief Financial Officer (Principal Financial and Accounting Officer) |
POWER OF ATTORNEY
Each person whose individual signature appears below hereby authorizes and appoints Steven D. Harr, M.D., Nathan Hardy, and James J. MacDonald and each of them, with full power of substitution and resubstitution and full power to act without the other, as his or her true and lawful attorney-in-fact and agent to act in his or her name, place and stead and to execute in the name and on behalf of each person, individually and in each capacity stated below, and to file any and all amendments to this Annual Report on Form 10-K and to file the same, with all exhibits thereto, and other documents in connection therewith, with the Securities and Exchange Commission, granting unto said attorneys-in-fact and agents, and each of them, full power and authority to do and perform each and every act and thing, ratifying and confirming all that said attorneys-in-fact and agents or any of them or their or his substitute or substitutes may lawfully do or cause to be done by virtue thereof.
Pursuant to the requirements of the Securities Exchange Act of 1934, as amended, this Annual Report on Form 10-K has been signed below by the following persons on behalf of the Registrant in the capacities and on the dates indicated.
|
Signature |
|
Title |
|
Date |
|
|
|
|
|
|
|
/s/ Steven D. Harr, M.D.
|
|
President, Chief Executive Officer and Director (Principal Executive Officer) |
|
March 16, 2022 |
|
Steven D. Harr, M.D. |
|
|
||
|
|
|
|
|
|
|
/s/ Nathan Hardy
|
|
Chief Financial Officer (Principal Financial and Accounting Officer) |
|
March 16, 2022 |
|
Nathan Hardy |
|
|
||
|
|
|
|
|
|
|
/s/ Hans E. Bishop
|
|
Chairman of the Board |
|
March 16, 2022 |
|
Hans E. Bishop |
|
|
||
|
|
|
|
|
|
|
/s/ Joshua H. Bilenker, M.D.
|
|
Director |
|
March 16, 2022 |
|
Joshua H. Bilenker, M.D. |
|
|
||
|
|
|
|
|
|
|
/s/ Douglas Cole, M.D.
|
|
Director |
|
March 16, 2022 |
|
Douglas Cole, M.D. |
|
|
||
|
|
|
|
|
|
|
/s/ Richard Mulligan, Ph.D.
|
|
Director |
|
March 16, 2022 |
|
Richard Mulligan, Ph.D. |
|
|
||
|
|
|
|
|
|
|
/s/ Robert Nelsen
|
|
Director |
|
March 16, 2022 |
|
Robert Nelsen |
|
|
||
|
|
|
|
|
|
|
/s/ Alise S. Reicin, M.D.
|
|
Director |
|
March 16, 2022 |
|
Alise S. Reicin, M.D. |
|
|
||
|
|
|
|
|
|
|
/s/ Michelle Seitz
|
|
Director |
|
March 16, 2022 |
|
Michelle Seitz |
|
|
187
|
Signature |
|
Title |
|
Date |
|
|
|
|
|
|
|
|
|
|
|
|
|
/s/ Mary Agnes (Maggie) Wilderotter
|
|
Director |
|
March 16, 2022 |
|
Mary Agnes (Maggie) Wilderotter |
|
|
||
|
|
|
|
|
|
|
/s/ Patrick Y. Yang, Ph.D.
|
|
Director |
|
March 16, 2022 |
|
Patrick Y. Yang, Ph.D. |
|
|
188